| Herrerasaurus Fossil range: Late Triassic, 228 Ma | |
|---|---|
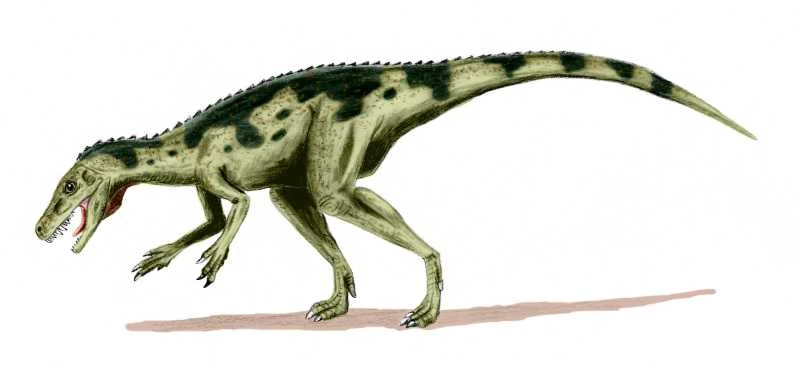 Artist's restoration of Herrerasaurus ischigualastensis. | |
| Scientific classification
| |
|
Herrerasaurus | |
| |
| |
Herrerasaurus (meaning "Herrera's lizard", after the name of the rancher who discovered the first fossil of the animal) was one of the earliest dinosaurs. All known specimens of this carnivore have been discovered in rocks of early Carnian age (late Triassic, around 228 million years ago) in northwestern Argentina. The type species, Herrerasaurus ischigualastensis, was described by Osvaldo Reig in 1963[1] and is the only species assigned to the genus. The names Ischisaurus and Frenguellisaurus are synonymous with Herrerasaurus.
For many years, the classification of Herrerasaurus was unclear, as the animal was initially known from very fragmentary remains; it has been hypothesized to be a basal theropod, a basal sauropodomorph, a basal saurischian, or not a dinosaur at all. However, with the discovery of a mostly complete skeleton and skull in 1988,[2][3] Herrerasaurus has been classified as either an early theropod or an early saurischian in at least five recent reviews of theropod evolution. Herrerasaurus is a medium-sized bipedal member of the Herrerasauridae, a group of similar animals which were among the earliest of the dinosaurian evolutionary radiation.
Description[]
{kind=link}
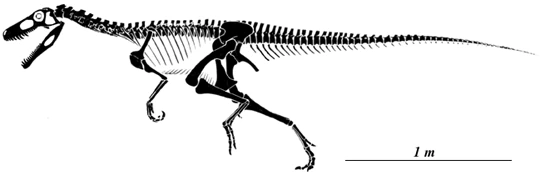
Skeletal reconstruction of Herrerasaurus.
Herrerasaurus was a lightly-built bipedal carnivore with a long tail and a relatively small head. Its length is estimated at 3 to 6 meters (10 to 20 ft),[3] and its hip height at more than 1.1 meters (3.3 ft).[4] It may have weighed around 210–350 kilograms (463–772 lb).[4] In a large specimen at first thought to belong to a separate genus, Frenguellisaurus, the skull measured 56 centimeters (1.8 ft) in length.[4] Smaller specimens had skulls which measured around 30 centimeters (1 ft) in length.[5]
Skull[]

{kind=link}
Mounted Herrerasaurus skeleton, at the Field Museum in Chicago.


{kind=link}
Skull of Herrerasaurus
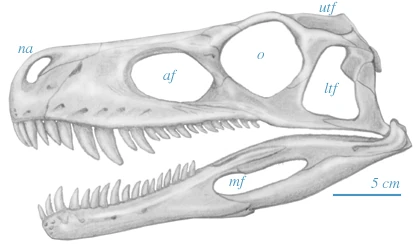

{kind=link}
Skull of Herrerasaurus ischigualastensis, based on Sereno and Novas (1993)
Herrerasaurus had a long, narrow skull that lacked nearly all the specializations that characterized later dinosaurs,[6] and more closely resembled those of more primitive archosaurs such as Euparkeria. It had five pairs of fenestrae (skull openings) in its skull, two pairs of which were for the eyes and nostrils. Between the eyes and the nostrils were two antorbital fenestrae and a pair of tiny, 1-centimeter-long (0.4 in) slit-like holes called promaxillary fenestrae.[7] Behind the eyes were large infratemporal fenestrae. These holes helped to reduce the weight of the skull.
Herrerasaurus had a flexible joint in the lower jaw, allowing it to slide back and forth to deliver a grasping bite.[6] This cranial specialization is unusual among the dinosaurs but has evolved independently in some lizards.[8] The rear of the lower jaw also had fenestrae. The jaws were equipped with large serrated teeth for biting and eating flesh, and the neck was slender and flexible.[6][9]
Limbs[]
Herrerasaurus had relatively short forelimbs, which were less than half the length of its hind limbs. The upper arm and forearm were rather short, while the manus was elongated. The first two fingers and the thumb bore curved, sharp claws for grasping prey. Its fourth and fifth digits were small stubs without claws.[3][10]Herrerasaurus was bipedal. It had strong hind limbs with short thighs and rather long feet, indicating this animal was most likely a swift runner. The balancing tail, partially stiffened by overlapping vertebral processes, also indicates an adaptation for speed.[5]
Unlike most reptiles of its era, Herrerasaurus was fully bipedal. It had strong hind limbs with short thighs and rather long feet, indicating that it was most likely a swift runner. The foot had five toes, but only the middle three (digits II, III, and IV) bore weight. The outer toes (I and V) were small; the first toe had a small claw. The tail, partially stiffened by overlapping vertebral projections, balanced the body and was also an adaptation for speed.[5]
Derived and basal characteristics[]
Herrerasaurus is something of an enigma in that it displays traits that are found in different groups of dinosaurs, and several traits found in non-dinosaurian archosaurs. Although it shared most of the characteristics of dinosaurs, there were a few differences, particularly in regard to the shape of its hip and leg bones. Its pelvis was similar to that of saurischian dinosaurs, but it had a bony acetabulum (where the femur meets the pelvis) that was only partially open. The ilium, the main hip bone, was supported only by two sacrals, a basal trait,[5] but the pubis pointed backwards, a derived trait that parallels what is seen in dromaeosaurids and birds. Additionally, the end of the pubis had a booted shape, similar to what is present in avetheropods, and the vertebral centra had an Allosaurus-like hourglass shape.[4]
Classification[]
|
Herrerasaurus gives its name to its family, Herrerasauridae, of the mid- to late Triassic, although where it and its close relatives lie on the early dinosaur evolutionary tree is unclear. They are possibly basal theropods or basal saurischians but may in fact predate the saurischian-ornithischian split.[12] The situation is further complicated by uncertainties in correlating the ages of late Triassic beds bearing land animals.[5] Other members of the clade may include Eoraptor from the same Ischigualasto Formation of Argentina as Herrerasaurus, Staurikosaurus from the Santa Maria Formation of southern Brazil,[13] Chindesaurus from the Upper Petrified Forest (Chinle Formation) of Arizona,[14] and possibly Caseosaurus from the Dockum Formation of Texas,[15] although the relationships of these animals are not fully understood, and not all paleontologists agree. Other possible basal theropods, Alwalkeria from the Late Triassic Maleri Formation of India,[16] and Teyuwasu, known from very fragmentary remains from the Late Triassic of Brazil, might be related.[17] Novas (1992) defined Herrerasauridae as Herrerasaurus, Staurikosaurus, and their most recent common ancestor.[18] Sereno (1998) defined the group as the most inclusive clade including H. ischigualastensis but not Passer domesticus.[19] Langer (2004) created a higher level taxon, infraorder Herrerasauria.[5]
History[]


{kind=link}
San Juan region of Argentina, Badlands of the Valle de la Luna, where Herrerasaurus remains were unearthed.


{kind=link}
Skeletal reconstruction of Herrerasaurus on display in Mexico.
Herrerasaurus was named by paleontologist Osvaldo Reig after Victorino Herrera, an Andean goatherd who first noticed its fossils in outcrops near the city of San Juan in 1959.[1] These rocks, which later yielded Eoraptor,[20] are part of the Ischigualasto Formation and date from the late Ladinian to early Carnian stages of the Late Triassic period.[21] Reig named a second dinosaur from these rocks in the same publication as Herrerasaurus;[1] this dinosaur, Ischisaurus cattoi, is now considered a junior synonym and a juvenile of Herrerasaurus.[9] Two other partial skeletons, with skull material, were named Frenguellisaurus ischigualastensis by Fernando Novas in 1986,[22] but this species too is now thought to be a synonym.[9]
Reig believed Herrerasaurus was an early example of a carnosaur,[1] but this was the subject of much debate over the next 30 years, and the genus was variously classified during that time. In 1970, Steel classified Herrerasaurus as a prosauropod.[23] In 1972, Peter Galton classified the genus as not diagnosable beyond Saurischia.[24] Later, using cladistic analysis, some researchers put Herrerasaurus and Staurikosaurus at the base of the dinosaur tree before the separation between ornithischians and saurischians.[12][18][25][26] Several researchers classified the remains as non-dinosaurian.[27]


{kind=link}
Herrerasaurus skeleton and restoration on display at the Chicago Field Museum of Natural History.
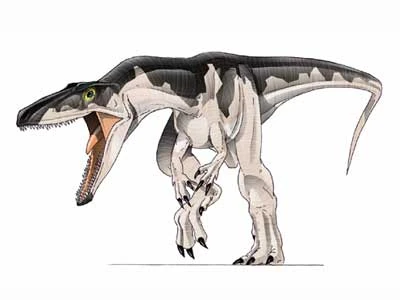

{kind=link}
Alternative restoration of Herrerasaurus.
A complete Herrerasaurus skull was not found until 1988, by a team of paleontologists led by Paul Sereno.[3] Based on the new fossils, authors such as Thomas Holtz[28] and Jose Bonaparte[29] classified Herrerasaurus at the base of the saurischian tree before the divergence between prosauropods and theropods. However, Sereno favored classifying Herrerasaurus (and the Herrerasauridae) as primitive theropods. These two classifications have become the most persistent, with Rauhut (2003)[30] and Bittencourt and Kellner (2004)[31] favoring the early theropod hypothesis, and Max Langer (2004),[5] Langer and Benton (2006),[32] and Randall Irmis and his coauthors (2007)[33] favoring the basal saurischian hypothesis. If Herrerasaurus were indeed a theropod, it would indicate that theropods, sauropodomorphs, and ornithischians diverged even earlier than herrerasaurids, before the middle Carnian, and that "all three lineages independently evolved several dinosaurian features, such as a more advanced ankle joint or an open acetabulum".[34] This view is further supported by ichnological records showing large tridactyl (three-toed) footprints that can be attributed only to a theropod dinosaur. These footprints date from the Ladinian (Middle Triassic) of the Los Rastros Formation in Argentina and predate Herrerasaurus by 3 to 5 million years.[35][36]
The study of early dinosaurs such as Herrerasaurus and Eoraptor therefore has important implications for the concept of dinosaurs as a monophyletic group (a group descended from a common ancestor). The monophyly of dinosaurs was explicitly proposed in the 1970s by Galton and Robert T. Bakker,[37][38] who compiled a list of cranial and postcranial synapomorphies (common anatomical traits derived from the common ancestor). Later authors proposed additional synapomorphies.[12][18] An extensive study of Herrerasaurus by Sereno in 1992 suggested that of these proposed synapomorphies, only one cranial and seven postcranial features were actually derived from a common ancestor, and that the others were attributable to convergent evolution. Sereno's analysis of Herrerasaurus also led him to propose several new dinosaurian synapomorphies.[3]
Paleoecology[]


{kind=link}
Herrerasaurus skeleton, North American Museum of Ancient Life.
Although Herrerasaurus shared the body shape of the large carnivorous dinosaurs, it lived about 230 million years ago, a time when dinosaurs were small and insignificant. It was the time of non-dinosaurian reptiles, not dinosaurs, and a major turning point in the Earth's ecology. The vertebrate fauna of the Ischigualasto Formation and the slightly later Los Colorados Formation consisted mainly of a variety of crurotarsal archosaurs and synapsids.[39][40] For instance, in the Ischigualasto Formation, dinosaurs constituted only about 6% of the total number of fossils.[41] By the end of the Triassic Period, dinosaurs were becoming the dominant large land animals, and the other archosaurs and synapsids declined in variety and number.[42]
Studies suggest that the paleoenvironment of the Ischigualasto Formation was a volcanically active floodplain covered by forests and subject to strong seasonal rainfalls. The climate was moist and warm,[43] though subject to seasonal variations.[44] Vegetation consisted of ferns (Cladophlebis), horsetails, and giant conifers (Protojuniperoxylon).[40] These plants formed highland forests along the banks of rivers.[3] Herrerasaurus remains appear to have been the most common among the carnivores of the Ischigualasto Formation.[21] It lived in the jungles of Late Triassic South America alongside another early dinosaur, the one metre long Eoraptor, as well as Saurosuchus,[45] a giant land-living rauisuchian (a quadrupedal meat eater with a theropod-like skull); the broadly similar but smaller Venaticosuchus, an ornithosuchid; and the predatory chiniquodontids.[21] Herbivores were much more abundant than carnivores and were represented by rhynchosaurs such as Hyperodapedon (a beaked reptile); aetosaurs (spiny armored reptiles); kannemeyeriid dicynodonts (stocky, front-heavy beaked quadrupedal animals) such as Ischigualastia; and traversodontids (somewhat similar in overall form to dicynodonts, but lacking beaks) such as Exaeretodon. These non-dinosaurian herbivores were much more abundant than early ornithischian dinosaurs like Pisanosaurus.[46]
Paleobiology[]
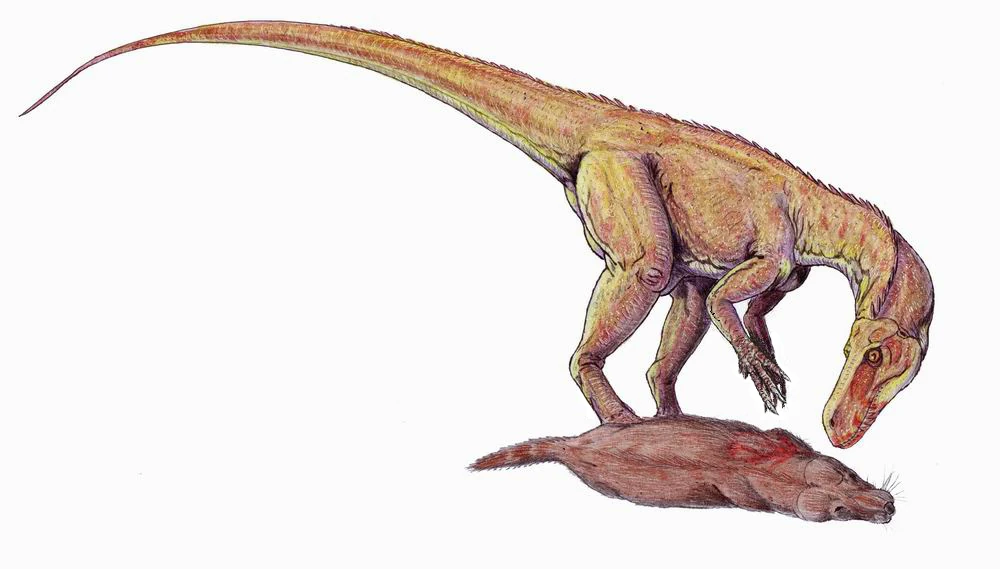
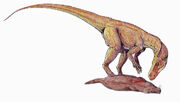
{kind=link}
Herrerasaurus feeding on a small synapsid
The teeth of Herrerasaurus indicate that it was a carnivore; its size indicates it would have preyed upon small and medium-sized plant eaters. These might have included other dinosaurs, such as Pisanosaurus, as well as the more plentiful rhynchosaurs and synapsids.[47] Herrerasaurus itself may have been preyed upon by giant rauisuchids like Saurosuchus; puncture wounds were found in one skull.[6]
Coprolites (fossilized dung) containing small bones but no trace of plant fragments, discovered in the Ischigualasto Formation, have been assigned to Herrerasaurus based on fossil abundance. Mineralogical and chemical analysis of these coprolites indicates that this carnivore could digest bone.[48]
Similar studies made on coprolites assigned to a large Cretaceous theropod (most probably Tyrannosaurus) and concluding to the ability of dinosaurs to dissolve ingested bones, further strengthen the assertion.[49] Karen's paper said the opposite, that they were passing the bones quickly without digestion.
References[]
- ^ a b c d Reig, O.A. (1963). "La presencia de dinosaurios saurisquios en los "Estratos de Ischigualasto" (Mesotriásico Superior) de las provincias de San Juan y La Rioja (República Argentina)" (in Spanish). Ameghiniana 3 (1): 3–20.
- ^ Sereno, P.C.; Novas, F.E.; Arcucci, A.B.; and C. Yu (1988). "New evidence on dinosaur and mammal origins from the Ischigualasto Formation (Upper Triassic, Argentina)". Journal of Vertebrate Paleontology 8 (3, supplement): 26A.
- ^ a b c d e f Sereno, P.C.; and Novas, F.E. (1992). "The complete skull and skeleton of an early dinosaur". Science 258: 1137–1140. doi:. PMID 17789086.
- ^ a b c d Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. pp. 248–250. ISBN 0671687336.
- ^ a b c d e f g h Langer, Max C. (2004). "Basal Saurischia". in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 25–46. ISBN 0-520-24209-2. Cite error: Invalid
<ref>tag; name "Langer04" defined multiple times with different content - ^ a b c d Sereno, P.C.; and Novas, F.E. (1993). "The skull and neck of the basal theropod Herrerasaurus ischigualastensis". Journal of Vertebrate Paleontology 13: 451–476.
- ^ Sereno, P.C. (2007). "The phylogenetic relationships of early dinosaurs: a comparative report". Historical Biology 19 (1): 145–155. doi:. ISSN 0891-2963.
- ^ McDowell, S.B., Jr.; and C.M. Bogert (1954). "The systematic position of Lanthanotus and the affinities of the anguinomorphan lizards". Bulletin of the American Museum of Natural History 105: 1–142.
- ^ a b c Novas, F. E. (1994). "New information on the systematics and postcranial skeleton of Herrerasaurus ischigualastensis (Theropoda: Herrerasauridae) from the Ischigualasto Formation (Upper Triassic) of Argentina". Journal of Vertebrate Paleontology 13 (4): 400–423.
- ^ Sereno, P.C. (1993). "The pectoral girdle and forelimb of the basal theropod Herrerasaurus ischigualastensis". Journal of Vertebrate Paleontology 13: 425–450.
- ^ Benton, Michael J. (2004). "Origin and relationships of Dinosauria". in Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.). The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 7–19. ISBN 0-520-24209-2.
- ^ a b c Brinkman, D.B.; and Sues, H.-D. (1987). "A staurikosaurid dinosaur from the Upper Triassic Ischigualasto Formation of Argentina and the relationships of the Staurikosauridae". Palaeontology 30: 493–503.
- ^ Colbert, E.H. (1970). "A saurischian dinosaur from the Triassic of Brazil". American Museum Novitates 2405: 1–39.
- ^ Long, R.A.; and Murry, P.A. (1995). "Late Triassic (Carnian and Norian) Tetrapods from the Southwestern United States". New Mexico Museum of Natural History and Science, Bulletin 4: 1–254.
- ^ Hunt, A.P.; Lucas, S.G.; Heckert, A.B.; Sullivan, R.M.; and Lockley, M.G. (1998). "Late Triassic Dinosaurs from the Western United States". Geobios 31 (4): 511–531. doi:.
- ^ Chatterjee, S.; and Creisler, B.S. (1994). "Alwalkeria (Theropoda) and Morturneria (Plesiosauria), new names for preoccupied Walkeria Chatterjee, 1987 and Turneria Chatterjee and Small, 1989". Journal of Vertebrate Paleontology (1): 142.
- ^ Kischlat, E.-E. (1999). "A new dinosaurian “rescued” from the Brazilian Triassic: Teyuwasu barberenai, new taxon". 1st Simp. Brasil. Pal. Vert, Paleontologia em Destaque 14 (26): 58.
- ^ a b c Novas, F. E. (1992). "Phylogenetic relationships of the basal dinosaurs, the Herrerasauridae". Palaeontology 35: 51–62.
- ^ Sereno, P.C. (1998). "A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210 (1): 41–83.
- ^ Sereno, P.C.; Forster, C.A.; Rogers, R.R.; and Monetta, A.M. (1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosaurs". Nature 361 (6407): 64–66. doi:.
- ^ a b c Rogers, R. R.; Swisher III, C.C.; Sereno, P.C.; Monetta, A.M.; Forster, C.A.; and Martinez, R.N. (1993). "The Ischigualasto tetrapod assemblage (Late Triassic, Argentina) and 40Ar/39Ar dating of dinosaur origins". Science 260: 794–797. doi:. PMID 17746113.
- ^ Novas, F. E. (1986). "Un probable terópodo (Saurischia) de la Formación Ischigualasto (Triásico superior), San Juan, Argentina" (in Spanish). IV Congreso Argentino de Paleontología y Bioestratigrafía, Mendoza, november 23-27 2: 1–6.
- ^ Steel, R. (1970). "Part 14. Saurischia. Handbuch der Paläoherpetologie/Encyclopedia of Paleoherpetology". Gustav Fischer Verlag, Stuttgart: 1–87.
- ^ Galton, P.M. (1973). "On the anatomy and relationships of Efraasia diagnostica (Huene) n.gen., a prosauropod dinosaur (Reptilia: Saurischia) from the Upper Triassic of Germany.". Paläontologische Zeitschrift 47 (3/4): 229–255.
- ^ Paul, G.S. (1984). "The segnosaurian dinosaurs: relics of the prosauropod-ornithischian transition?". Journal of Vertebrate Paleontology 4: 507–515.
- ^ Novas, F. E. (1989). "The tibia and tarsus in Herrerasauridae (Dinosauria, incertae sedis) and the evolution and origin of the dinosaurian tarsus". Journal of Paleontology 63: 677–690.
- ^ Gauthier, J.A., Cannatella, D., Queiroz, K., Kluge, A.G., and Rowe, T. (1989). "Tetrapod phylogeny". in Fernholm, K. Bremer, & Jörnvall, H.. The Hierarchy of Life. Amsterdam: Elsevier Science Publishers B. V.. pp. 337–353.
- ^ Holtz, T.R., Jr.; and Padian, K. (1995). "Definition and diagnosis of Theropoda and related taxa". Journal of Vertebrate Paleontolology 15 (3, supplement): 35A.
- ^ Bonaparte, J.F.; and Pumares, J.A. (1995). "Notas sobre el primer cráneo de Riojasaurus incertus (Dinosauria, Prosauropoda, Melanorosauridae) del Triásico superior de La Rioja, Argentina" (in Spanish). Ameghiniana 32: 341–349.
- ^ Rauhut, O.W.M. (2003). "The interrelationships and evolution of basal theropod dinosaurs". Special Papers in Palaeontology 69: 1–213.
- ^ Bittencourt, J.; and Kellner, W.A. (2004). "The phylogenetic position of Staurikosaurus pricei from the Triassic of Brazil". Journal of Vertebrate Paleontology 24 (3, supplement): 39A.
- ^ Langer, M.C; and Benton, M.J. (2006). "Early dinosaurs: a phylogenetic study". Journal of Systematic Palaeontology 4 (4): 309–358. doi:.
- ^ Irmis, Randall B.; Nesbitt, Sterling J.; Padian, Kevin; Smith, Nathan D.; Turner, Alan H.; Woody, Daniel; and Downs, Alex (2007). "A Late Triassic dinosauromorph assemblage from New Mexico and the rise of dinosaurs". Science 317: 358–361. doi:. PMID 17641198.
- ^ White, A.T.; and Kazlev, M.A. (2003). "Theropoda: Basal Theropods". Palaeos.com. http://www.palaeos.com/Vertebrates/Units/340Theropoda/340.100.html. Retrieved on 2007-07-20.
- ^ Forster, C.A.; Arcucci, A.B.; Marsicano, C.A.; Abdala, F.; and May, C.L. (1995). "New vertebrate material from the Los Rastros Formation (Middle Triassic), La Rioja province, northwestern Argentina". Journal of Vertebrate Paleontology 15 (3, supplement): 29A.
- ^ Arcucci, A. C.; Forster, C.; May, Abdala, F.; and Marsicano, C. (1998). "Una nueva icnofauna de la Formación Los Rastros, Triásico Medio, en la Quebrada de Ischichuca Chica (Provincia de La Rioja, Argentina)" (in Spanish). Acta Geológica Lilloana 18: 152.
- ^ Bakker, R.T.; and Galton, P.M. (1974). "Dinosaur monophyly and a new class of vertebrates". Nature 248: 168–172. doi:.
- ^ Bonaparte, J. F. (1976). "Pisanosaurus mertii Casimiquela and the origin of the Ornithischia". Journal of Paleontology 50: 808–820..
- ^ Bonaparte, J. F. (1982). "Faunal replacement in the Triassic of South America". Journal of Vertebrate Paleontology 2: 362–371.
- ^ a b Bonaparte, J.F. (1979). "Faunas y paleobiogeografía de los tetrápodos mesozoicos de América del Sur" (in Spanish). Ameghiniana, Revista de la Asociación Paleontológica Argentina 16 (3–4): 217–238.
- ^ Benton, Michael J. (1999). "Origin and early evolution of dinosaurs". in Farlow, James O.; Brett-Surman, M.K.. The Complete Dinosaur. Indiana University Press. pp. 204–215. ISBN 0-253-21313-4.
- ^ Parrish, J. Michael (1999). "Evolution of the archosaurs". in Farlow, James O.; Brett-Surman, M.K.. The Complete Dinosaur. Indiana University Press. pp. 191–203. ISBN 0-253-21313-4.
- ^ Tucker, Maurice E.; Benton, Michael J. (1982). "Triassic environments, climates, and reptile evolution". Palaeogeography, Palaeoclirnatology, Palaeoecology 40: 361–379. doi:. http://palaeo.gly.bris.ac.uk/benton/reprints/1982triassic.pdf. Retrieved on 2009-07-23.
- ^ Columbi, Carina E. (2008-10-05). "Stable isotope analysis of fossil plants from the Upper Triassic Ischigualasto Formation in the northwest of Argentina"., Houston, TX: The Geological Society of America. Retrieved on 2009-07-23.
- ^ Sill, W.D. (1974). "The anatomy of Saurosuchus galilei and the relationships of the rauisuchid thecodonts". Bulletin of the Museum of Comparative Zoology 146: 317–362.
- ^ Bonaparte, J.F. (1970). "Annotated list of the South American Triassic tetrapods". Gondwana Symposium Proceedings and Papers 2: 665–682.
- ^ Sues, H.D. (1990). "Staurikosaurus and Herrerasauridae". in Weishampel, D.B.; Dodson, P.; Osmólska, H.. The Dinosauria. University of California Press. pp. 143–47. ISBN 0-520-06726-6.
- ^ Hollocher, K.T.; Alcober, O.A.; Colombi, C.E.; and Hollocher, T.C. (2005). "Carnivore coprolites from the Upper Triassic Ischigualasto Formation, Argentina: chemistry, mineralogy, and evidence for rapid initial mineralization". PALAIOS 20: 51–63. doi:.
- ^ Chin, K.; Tokaryk, T.T., Erickson, G.M., and Calk, L.C. (1998). "A king-sized theropod coprolite". Nature 393: 680–682. doi:10.1038/31461.
External links[]
- Introduction to Herrerasaurus, from the University of California Museum of Paleontology
- Herrerasaurus ischigualastensis, DinoData.org