| Temnospondyli Fossil range: Mississippian–Early Cretaceous | |
|---|---|
 Capetus palustris, a basal temnospondyl. | |
| Scientific classification
| |
|
Temnospondyli | |
|
See below. | |

{kind=link}
Koskinonodon, American Museum of Natural History
Temnospondyli are an important and extremely diverse taxon of small to giant primitive amphibians that flourished worldwide during the Carboniferous, Permian, and Triassic periods. At the end of the Triassic most temnospondyls vanished from the fossil record, but a few aquatic forms persisted in East Asia and Australia through the Jurassic, and a few stragglers continued into the Early Cretaceous.
Temnospondyli are one of the largest recognized groups of early amphibians with about 180 described genera. The Permian-Triassic extinction event wiped out several families, but the surviving lineages rediversified, and temnospondyls remained numerous through the Triassic before declining in the later Mesozoic era, the last survivor (Koolasuchus) is known from fossils found in the mid-Cretaceous rocks of Australia.
Most temnospondyls were 30 cm (1 ft) to 1 m (3.3 ft) in length, with a few forms growing to 2 m (6 ft), and one example, Prionosuchus, reaching lengths of 30 feet. During their evolutionary history they adapted to a very wide range of habitats, including fresh-water aquatic, semi-aquatic, amphibious, terrestrial, and in one group even near-shore marine, and their fossil remains have been found on every continent. Authorities disagree over whether some specialized forms were ancestral to some modern amphibians, or whether the whole group died out without leaving any descendants.[1][2][3]
Description[]


{kind=link}
Skeleton of Eryops megacephalus.
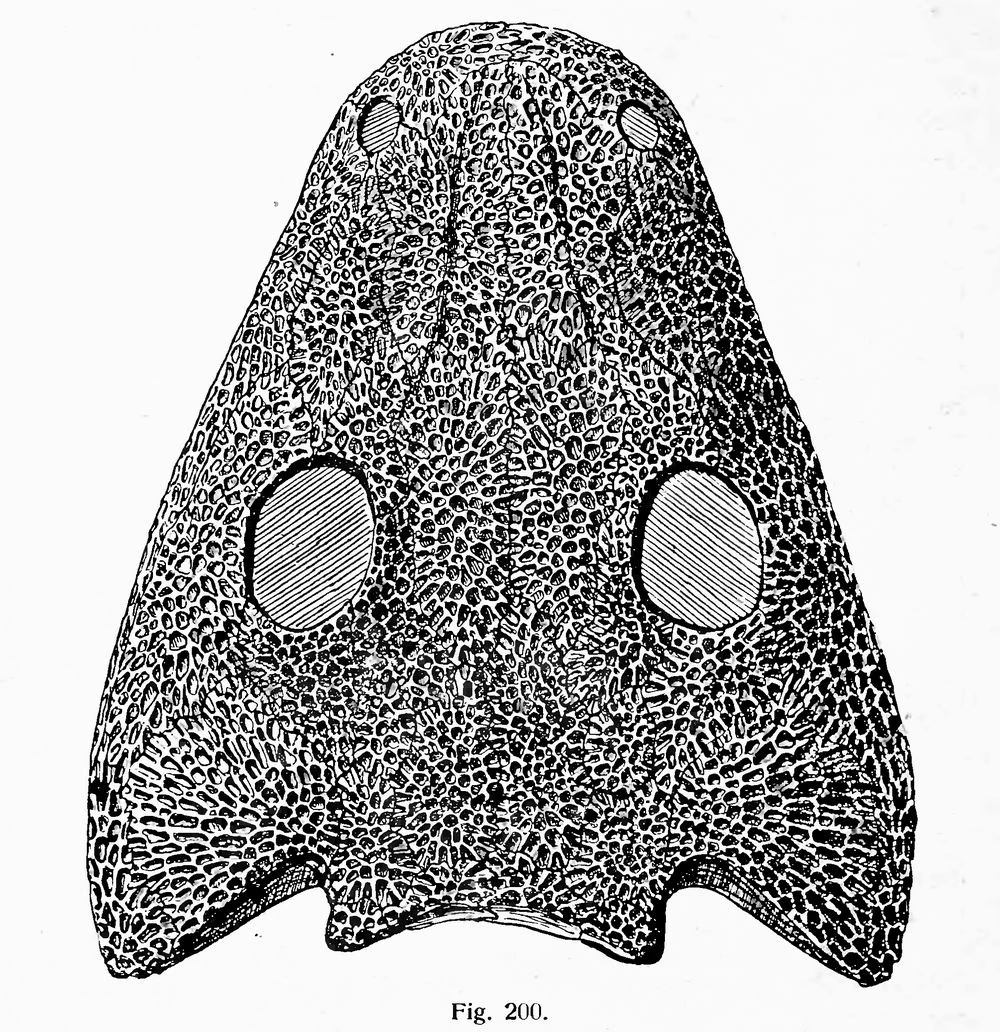

{kind=link}
Skull of Chelyderpeton
Early temnospondyls were similar to modern-day salamanders with large flat heads, no necks, short limbs, and an undulating gait. In the context of other early amphibians, they are recognized by the large openings across the roof of the mouth (palatal vacuities) and by the semicircular embayments behind the cheek region for froglike large eardrums.[4] They had short straight ribs which combined with the large skulls, suggest that they breathed by buccal pumping, ramming air into the lungs, and not using costal respiration.[5] The individual vertebrae were of multipart construction with the body (centrum) made up of three separate bones, namely one large intercentrum anteriorly and two smaller pleurocentra posteriorly. The skull was massively constructed and survives intact in the fossil record even when the rest of the skeleton is lost.[6]
Evolutionary History[]
Carboniferous temnospondyls[]


-RedpathMuseumMontreal-June6-08.png){kind=link}
A fossil specimen of the skull of Dendrerpeton, a temnospondyl from the Carboniferous.
During the Carboniferous, Temnospondyls included primitive medium-sized (Dendrerpeton) or large (Cochleosaurus about 1.5 meters long) semi-aquatic forms. Others, more advanced, were small and resembled newts or salamanders (Limnerpeton), and one group, represented by the genus Branchiosaurus and its relatives, even retained external gills like the modern-day axolotl. During the Late Carboniferous and Early Permian, several groups evolved strong, robust limbs and vertebrae, and became adapted to life on land (Cacops) or as large (c. 1.5 meters long) and heavy-bodied semi-aquatic predators (Trematops, Eryops). Others developed long snouts and an astonishing similarity to crocodiles (e.g. Archegosaurus) although lacking armour. This last group included the largest known amphibian, the 9 meter long Prionosuchus of Brazil.
Temnospondyls in the Permian[]
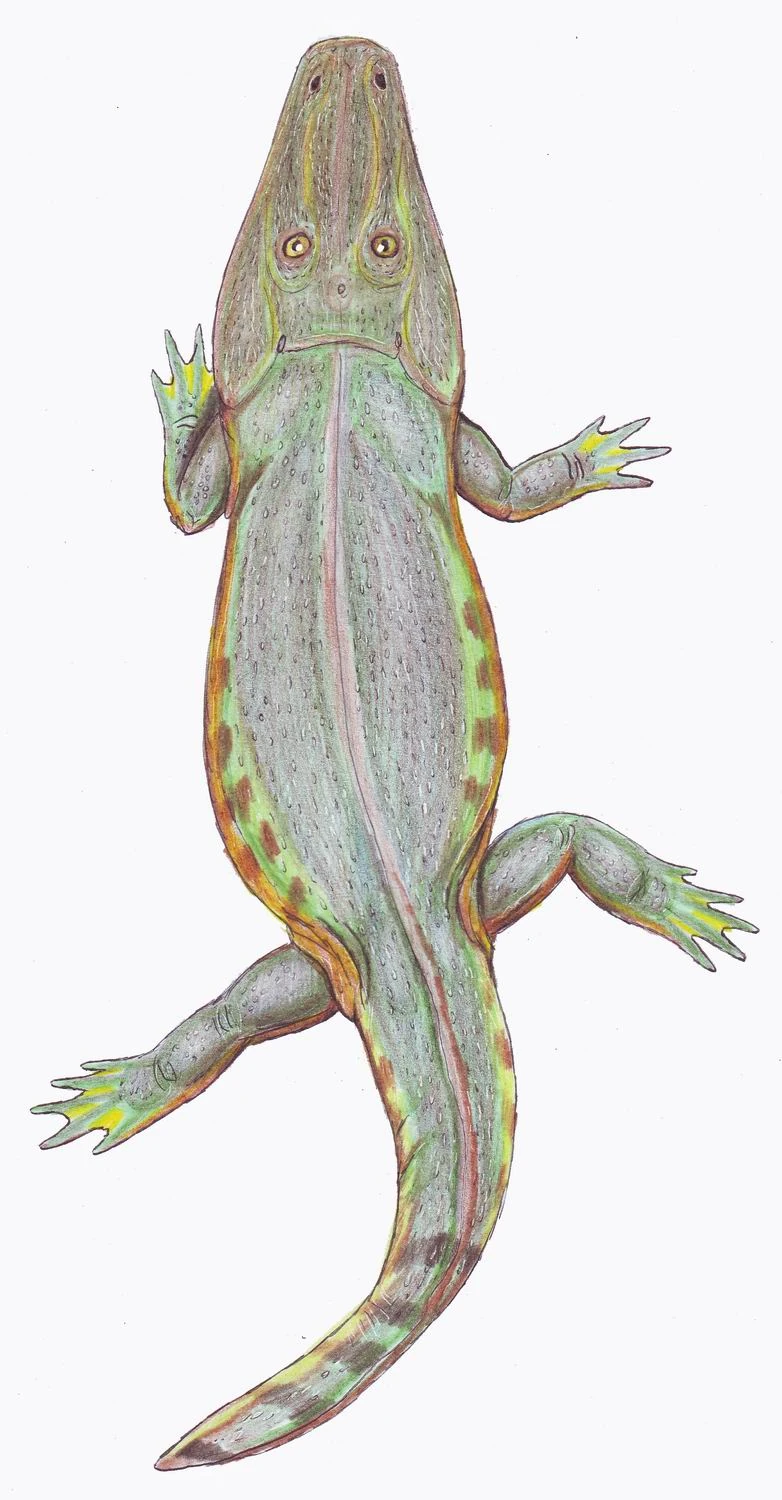

{kind=link}
Tryphosuchus, a Permian temnospondyl.
During the later Permian, increasing aridity and more successful reptiles meant the end of the terrestrial forms, but semi- and fully aquatic animals continued to flourish, including the large Melosaurus of Eastern Europe.
As these amphibians continued to flourish and diversify in the lakes and rivers of the Late Permian, a number of groups became more dependent on life in the water. The vertebrae became weak, the limbs small and vestigal, and the heavy skull large and flat, with the eyes looking upwards. After the Permian-Triassic extinction event, one subgroup of temnospondyls, the Stereospondyli, produced a new adaptive radiation of crocodile-like forms, mostly amphibious or aquatic and growing to 5 m (16.5 ft), such as Mastodonsaurus. During the Triassic period these animals dominated the fresh-water ecosystems, evolving in a range of both small and large forms. In some of these families, the vertebrae become further modified, with the main body of the vertebrae being reduced to a single large intercentrum. Many of the families forming this radiation were global in distribution. During the Early Triassic one group of successful long-snouted fish eaters, the Trematosaurs, even adapted to a life in the sea, the only known amphibians to do so (except the modern Crab-eating Frog).
The Capitosauroidea included not only medium-sized but also many giant species, 2.3 to 4 meters or more in length (e.g. Paracyclotosaurus, Cyclotosaurus), with huge and extraordinarily flat skulls, over a meter long in the largest forms (Mastodonsaurus). These animals seem to have lived on the river bottom, perhaps spending most or all their entire lives in water, and catching their prey by a sudden opening of the upper jaw, sucking in any unwary fish or smaller tetrapod that happened to swim past. In the Late Triassic (Carnian) these big amphibains were joined by the superficially very similar Metoposauridae (1.5 meters long—and distinguished mainly by the different position of the eye-sockets), and the curious wide-headed Plagiosaurus (about a meter in length), with external gills.
Mesozoic survivors[]
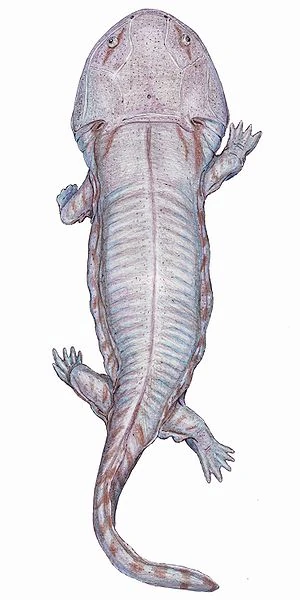
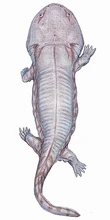
{kind=link}
Siderops kehli, a giant brachyopid from the Early Jurassic of Australia.
In the Late Triassic (Carnian), these big amphibains were joined by the superficially very similar Metoposauridae (1.5 meters long—and distinguished mainly by the different position of the eye-sockets), and the curious wide-headed Plagiosaurs (about a meter in length), with external gills.
The Triassic-Jurassic extinction event killed all the giant temnospondyls. Only the two Gondwanan families survived; the Brachyopidae and Chigutisauridae. Interestingly, these grew to large size during the Jurassic, with the Brachyopids flourishing in China, and the Chigutisaurs in Gondwana.
The most recent known temnospondyl was the giant Chigutisaur, Koolasuchus, known from the middle Cretaceous of Australia (where it seems to have survived in rift valleys that were too cold in the winter for crocodiles, co-existing with dinosaurs). At around 2.5 to 5 meters in length[7], this was one of the largest of its kind, as well as the last.
Forefathers of modern amphibians?[]
The Lissamphibia, the modern amphibians, appear to have risen in the Permian.[8] The root of the group is controversial, but may be with the Temnospondyli. Skull morphology of some of the smaller later form has been compared to those of modern frogs and salamanders. The presence of bicuspid, pedicellate teeth in small, paedomorphic or immature temnospondyls like the genus Doleserpeton has been cited as the most convincing argument in favor of the temnospondyl origin of lissamphibians.[9] Other analysis indicate that another fossil group, the Lepospondyli are more likely candidates for lissamphibian origin, leaving the Temnospondyli an extinct group.[10]
Systematics and phylogeny[]
Two types of vertebrae[]


{kind=link}
Originally, Temnospondyls were classified according to the structure of the vertebrae. Earlier forms, with complex vertebrae consisting of a number of separate elements, were called "Rachitomi", and large Triassic aquatic forms with simpler weaker vertebrae were called "Stereospondyli"; and these two types were considered suborders of the order Temnospondyli.
In the Rachitomous condition the intercentra were large and wedge-shaped, and the pleurocentra were relatively small blocks that fitted between them. Both elements supported the neural arch, and well-developed interlocking zygapophyses strengthened the connections between the vertebrae. The strong backbone and strong limbs let many ratchitomes be at least partially, and in some cases fully, terrestrial.
In the stereospondylous condition the pleurocentra have been lost, and the vertebral centra reduced to simple blocks made up of the intercentra only. This weaker type of backbone indicates a more fully aquatic existence.[11]
More recent research has shown that this classification is no longer viable. The basic rachitomous condition is primitive for amphibians (tetrapods) in general. The pure stereospondyls seem to have arisen from different ancestors.[12] Some temnospondyls have rachitomous, semi-rachitomous, and sterospondylous vertebrae at different points the vertebral column of the same individual. Other taxa have intermediate morphologies that do not fit into one or the other category.[13] However, at least according to some analyses, the stereospondyls (minus some ambiguous taxa) can still be considered a clade.[14][13]
Taxonomy[]
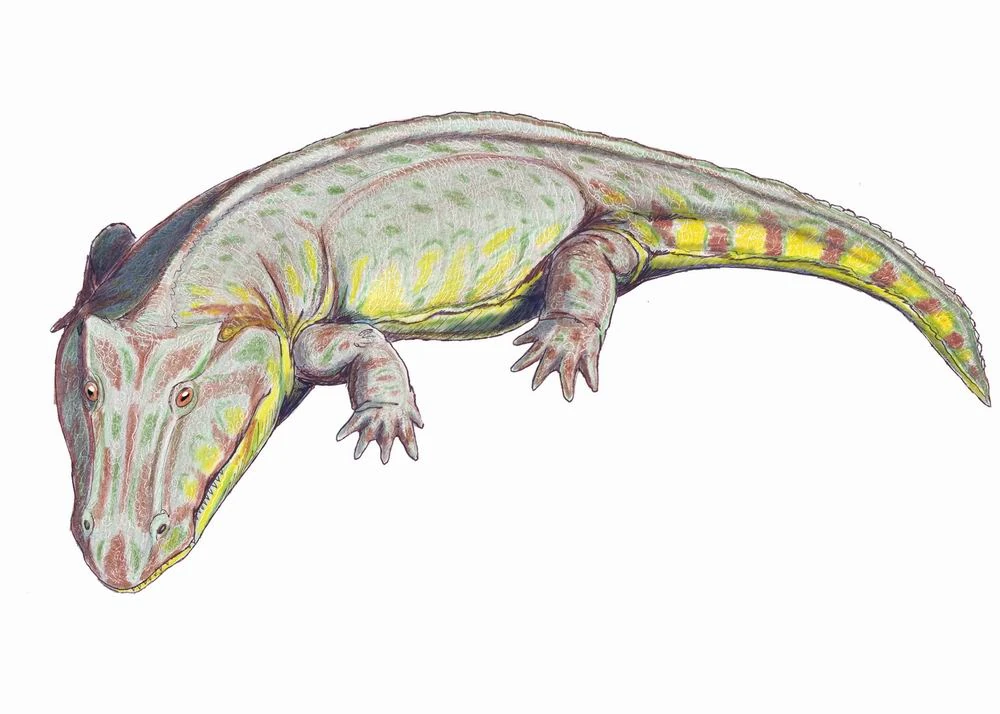
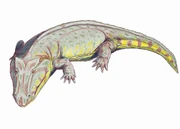
{kind=link}
Edops craigi, a basal edopoid.
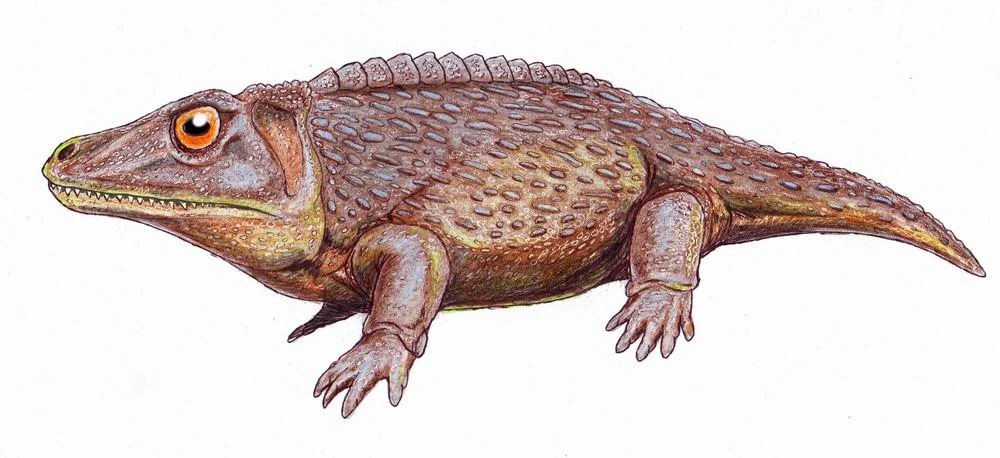

{kind=link}
Zygosaurus lucius, a dissorophid.
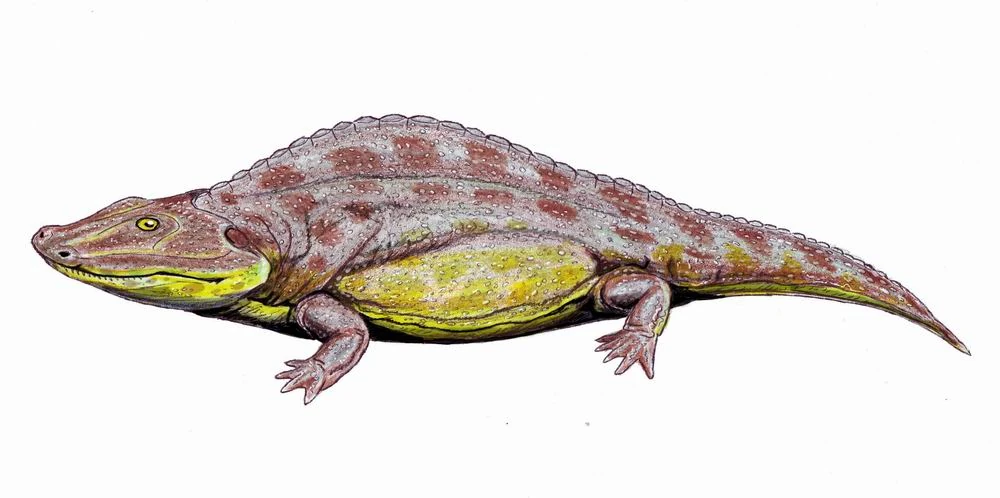

{kind=link}
Sclerothorax, a basal limnarchian.
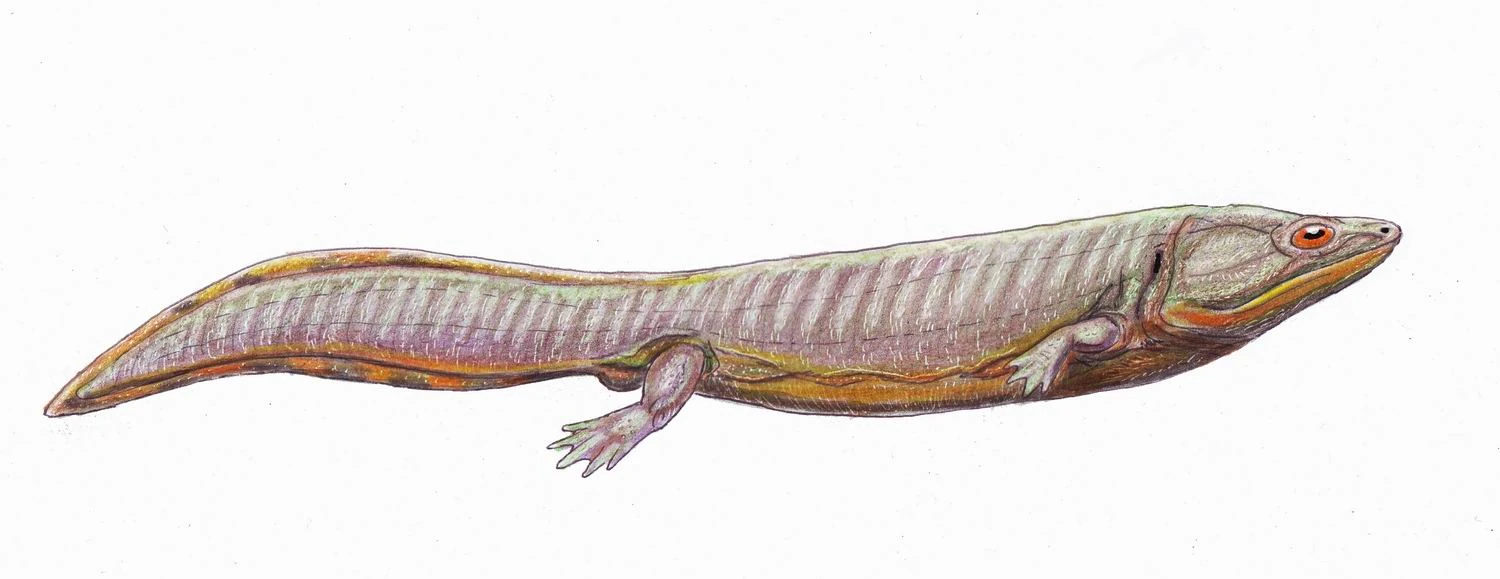

{kind=link}
Dvinosaurus egregius, a dvinosaurid.
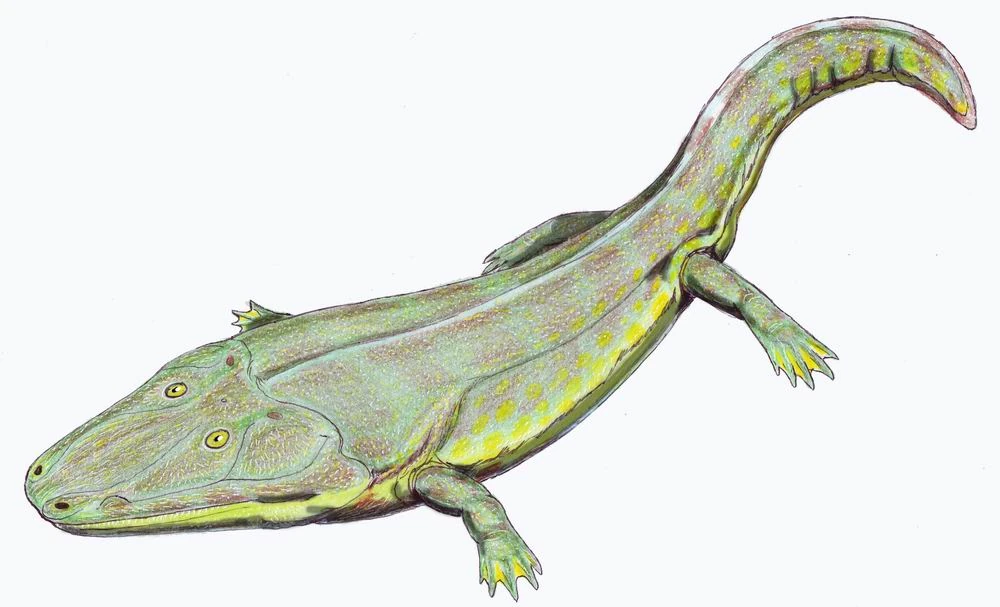

{kind=link}
Cyclotosaurus, a capitosaurid.
Class Amphibia
- Order Temnospondyli
- Saharastega
- Superfamily Edopoidea
- Family Cochleosauridae (Chenoprosopidae)
- Family Edopidae
- Capetus
- Iberospondylus
- Family Colosteidae
- Family Dendrerpetontidae
- Superfamily Capitosauroidea
- Suborder Euskelia
- Clade Limnarchia
- Suborder Dvinosauria
- Clade Stereospondylomorpha
- Superfamily Archegosauroidea
- Family Actinodontidae
- Family Archegosauridae
- Family Intasuchidae?
- Suborder Stereospondyli
- Family Peltobatrachidae
- Family Lapillopsidae
- Family Rhinesuchidae
- Family Lydekkerinidae
- Family Sclerothoracidae[15]
- Infraorder Trematosauria
- Superfamily Trematosauroidea
- Superfamily Metoposauroidea
- Superfamily Plagiosauroidea
- Superfamily Brachyopoidea
- SuperFamily Rhytidosteoidea
- Superfamily Archegosauroidea
Phylogeny[]
Studies on temnospondyl phylogeny mostly agree that 'post-edopoid' temnospondyls form a clade, the most basal members of which include Capetus, Dendrerpeton and Balanerpeton[16], although some paleontologists have found some of these taxa to be more basal than edopoids[17].
Among these basal 'post-edopoids', Dendrerpeton (from the Upper Carboniferous of Nova Scotia and Ireland) had a rather large skull with laterally facing orbits, a short body, and well-developed, robust limbs.[18] It lacked lateral line canals and grew to about 1 meter in length. These features suggest that it was predominantly terrestrial.[18]
Several species have been named.[19] Studies of a well-preserved, three-dimensional ear indicate that Dendrerpeton had a frog-like tympanum (ear drum) suited for the perception of airborne sounds.[20] This discovery provides support for the idea that lissamphibians descend from temnospondyls.
The following cladogram follows the analyses of Holmes, Carroll & Reisz (1998), Yates & Warren (2000), and Sidor et al. (2005).[21][14][22]
| Temnospondyli |
| |||||||||||||||||||||||||||||||||||||||||||||
References[]
- ^ Benton, M. J. (2000), Vertebrate Paleontology, 2nd Ed. Blackwell Science Ltd 3rd ed. (2004) - see also taxonomic hierarchy of the vertebrates, according to Benton 2004
- ^ Laurin, M. (1996) Terrestrial Vertebrates - Stegocephalians: Tetrapods and other digit-bearing vertebrates, The Tree of Life Web Project
- ^ Reisz, Robert, (no date), Biology 356 - Major Features of Vertebrate Evolution - The Origin of Tetrapods and Temnospondyls
- ^ R. Damiani, A systematic revision and phylogenetic analysis of Triassic mastodonsauroids (Temnospondyli, Stereospondyli), Zool. J. Linn. Soc., 133:379–482, 2001
- ^ H. Heatwole and R. L. Carroll (eds.), Amphibian Biology, vol. 4, Surrey Beatty, Chipping Norton, NSW, Australia, 2000
- ^ R. R. Schoch and A. R. Milner, Stereospondyli, vol. 33 of Encyclopedia of Palaeoherpetology, ed. by P. Wellnhofer, Pfeil, Munich, 2000
- ^ Adam Yates, Guide to Wild Dinosaurs, Sterling, 2003, p. 230 gives the length as 2.5 meters (8ft). According to Dann's Dinosaurs the total length is estimated at 4 to 5 metres (13-16 feet). Walking with Dinosaurs and Oz Fossils both give the length as 5 meters
- ^ Anderson J.S., Reisz R.R., Scott D., Fröbisch N.B., & Sumida S.S. (2008): A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders. Nature no 453: pp 515-518
- ^ Bolt, J. R. (1969): Lissamphibian origins: possible protolissamphibian from the Lower Permian of Oklahoma. Scienc no 166: pp 888-891
- ^ Laurin, M. (1998): The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I — systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: pp 1-42.
- ^ Colbert, E. H. (1969). Evolution of the Vertebrates, John Wiley & Sons Inc (2nd ed.)
- ^ Carroll, R. L. (1988), Vertebrate Paleontology and Evolution, W H Freeman & Co.
- ^ a b Laurin, M. and Steyer, J-S (2000) [* Laurin, M. and Steyer, J-S (2000) Phylogeny and Apomorphies of Temnospondyls, The Tree of Life Web Project
- ^ a b Yates, A. M. & Warren, A. A. (2000), The phylogeny of the 'higher' temnospondyls (Vertebrata: Choanata) and its implications for the monophyly and origins of the Stereospondyli. Zoological Journal of the Linnean Society 128: 77-121.
- ^ Schoch, R.R., Fastnacht, M., Fichter, J., and Keller, T. 2007. Anatomy and relationship of the Triassic temnospondyl Sclerothorax. Acta Palaeontologica Polonica 52, 117–136. (PDF)
- ^ (Milner & Sequeira 1994, 1998, Holmes et al. 1998, Ruta et al. 2003a, b)
- ^ (Steyer et al. 2006)
- ^ a b (Holmes et al. 1998)
- ^ (Milner 1996)
- ^ (Robinson et al. 2005)
- ^ Holmes, R.B., Carroll, R.L., and Reisz, R.R. (1998). "The first articulated skeleton of Dendrerpeton acadianum (Temnospondyli: Dendrerpentonidae) from the Lower Pennsylvanian locality of Joggins, Nova Scotia, and a review of its relationships." Journal of Vertebrate Paleontology, 18(1): 64-79.
- ^ Sidor, C.A., O'Keefe, F.R., Damiani, R.J., Steyer, J.-S., Smith, R.M.H., Larsson, H.C.E., Sereno, P.C., Ide, O., and Maga, A. (2005). "Permian tetrapods from the Sahara show climate-controlled endemism in Pangaea." Nature, 434: 886-889.
Further Reading[]
- Falcon-Lang, H. J. 1999. Fire ecology of a Late Carboniferous floodplain, Joggins, Nova Scotia. Journal of the Geological Society, London 156, 137-148.
- Holmes, R. B., Carroll, R. L. & Reisz, R. R. 1998. The first articulated skeleton of Dendrerpeton acadianum (Temnospondyli, Dendrerpetontidae) from the lower Pennsylvanian locality of Joggins, Nova Scotia, and a review of its relationships. Journal of Vertebrate Paleontology 18, 64-79.
- Milner, A. R. 1996. A revision of the temnospondyl amphibians from the Upper Carboniferous of Joggins, Nova Scotia. Special Papers in Palaeontology 52, 81-103.
- Sequeira, S. E. K. 1994. The temnospondyl amphibians from the Visean of East Kirkton, West Lothian, Scotland. Transactions of the Royal Society of Edinburgh: Earth Sciences 84, 331-361.
- Sequeira, S. E. K. 1998. A cochleosaurid temnospondyl amphibian from the Middle Pennsylvanian of Linton, Ohio, U.S.A. Zoological Journal of the Linnean Society 122, 261-290.
- Ruta, M., Coates, M. I. & Quicke, D. L. J. 2003a. Early tetrapod relationships revisited. Biological Reviews 78, 251-345.
External links[]
- Temnospondyli - Tree of Life project
- Temnospondyli - Palaeos
- Temnospondyli - Permian Tetrapods