{kind=link}
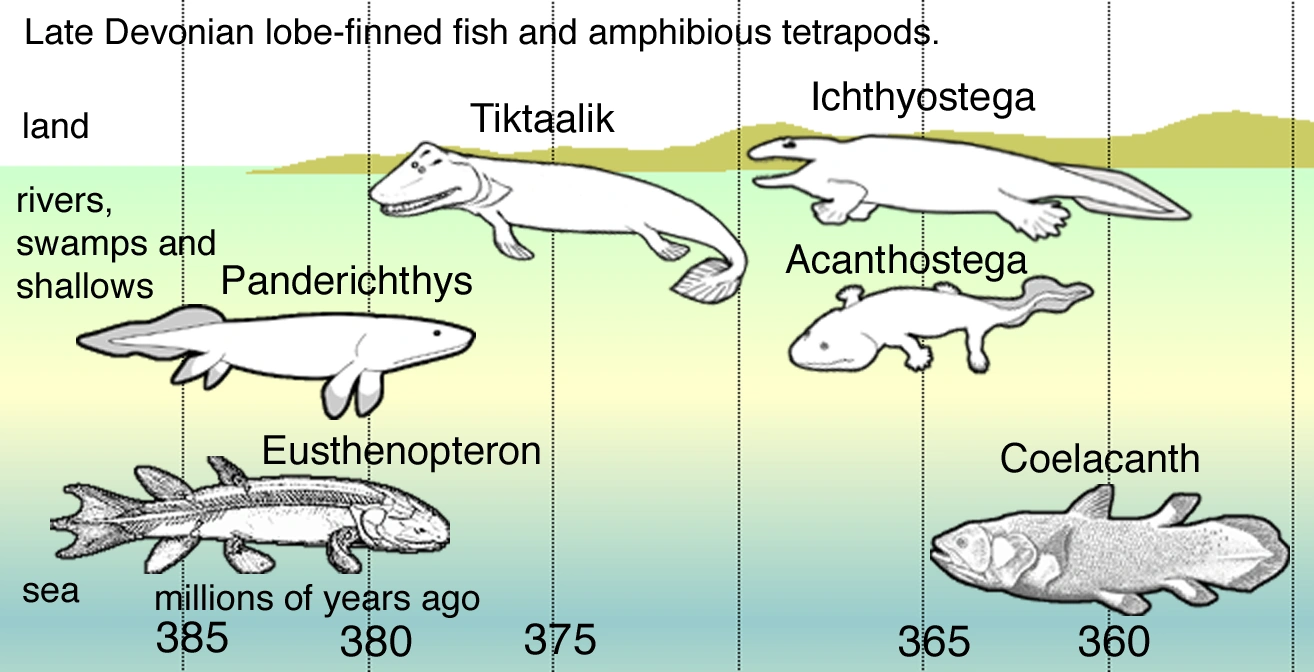
In Late Devonian vertebrate speciation, descendants of pelagic lobe-finned fish — like Eusthenopteron — exhibited a sequence of adaptations:
•Panderichthys, suited to muddy shallows;
•Tiktaalik with limb-like fins that could take it onto land;
•Early tetrapods in weed-filled swamps, such as;
•Acanthostega, which had feet with eight digits,
•Ichthyostega with limbs.
Descendants also included pelagic lobe-finned fish such as coelacanth species.
- This article primarily focuses on extinct tetrapods. For Wikipedia's article on Tetrapods, including modern-day ones, click here.
The earliest tetrapods radiated from the Sarcopterygii, or lobe-finned fish, evolving into air-breathing amphibians in the Devonian period.
Evolution[]

{kind=link}
Illustration showing the evolution of tetrapods from lobe-finned fish.
Devonian tetrapods[]

{kind=link}


{kind=link}


{kind=link}


{kind=link}


{kind=link}


{kind=link}

{kind=link}
Research by Jennifer A. Clack and her colleagues showed that the earliest tetrapods, such as Acanthostega, were wholly aquatic and quite unsuited to life on land. This overturned the earlier view that fish had first invaded the land - either in search of prey (like modern mudskippers) or to find water when the pond they lived in dried out - and later evolved legs, lungs, etc.
The first tetrapods are now thought to have evolved in shallow and swampy freshwater habitats, towards the end of the Devonian, a little more than 360 million years ago. By the late Devonian, land plants had stabilized freshwater habitats, allowing the first wetland ecosystems to develop, with increasingly complex food webs that afforded new opportunities. [1] Freshwater habitats were not the only places to find water filled with organic matter and choked with plants with dense vegetation near the water's edge. Swampy habitats like shallow wetlands, coastal lagoons and large brackish river deltas also existed at this time, and there is much to suggest that this is the kind of environment in which the tetrapods evolved. Early fossil tetrapods have been found in marine sediments, and because fossils of primitive tetrapods in general are found scattered all around the world, they must have spread by following the coastal lines—they could not have lived in freshwater only.
The common ancestor of all present gnathostomes lived in freshwater, and later migrated back to the sea. To deal with the much higher salinity in sea water, they evolved the ability to turn the nitrogen waste product ammonia into harmless urea, storing it in the body to make the blood as salty as the sea water without poisoning the organism. Ray-finned fishes later returned to freshwater and lost this ability. Since their blood contained more salt than freshwater, they could simply get rid of ammonia through their gills. When they finally returned to the sea again, they could not recover their old trick of turning ammonia to urea, and they had to evolve salt excreting glands instead. Lungfishes do the same when they are living in water, making ammonia and no urea, but when the water dries up and they are forced to burrow down in the mud, they switch to urea production. Like cartilaginous fishes, the coelacanth can store urea in its blood, as can the only known amphibians which can live for long periods of time in salt water (the toad Bufo marinus and the frog Rana cancrivora). These are traits they have inherited from their ancestors.
If early tetrapods lived in freshwater, and if they lost the ability to produce urea and used ammonia only, they would have to evolve it from scratch again later. Not a single species of all the ray-finned fishes living today has been able to do that, so it is not likely the tetrapods would have done so either. Terrestrial animals that can only produce ammonia would have to drink constantly, making a life on land impossible (a few exceptions exist, as some terrestrial woodlice can excrete their nitrogenous waste as ammonia gas). This probably also was a problem at the start when the tetrapods started to spend time out of water, but eventually the urea system would dominate completely. Because of this it is not likely they emerged in freshwater (unless they first migrated into freshwater habitats and then migrated onto land so shortly after that they still hadn't forgot how to make urea), even if some who never went to land (or extinct primitive species that returned to water) of course could have adapted to freshwater lakes and rivers.
Primitive tetrapods developed from a lobe-finned fish (an "osteolepid Sarcopterygian"), with a two-lobed brain in a flattened skull, a wide mouth and a short snout, whose upward-facing eyes show that it was a bottom-dweller, and which had already developed adaptations of fins with fleshy bases and bones (the "living fossil" coelacanth is a related marine lobe-finned fish without these shallow-water adaptations).
Even closer related was Panderichthys, who even had a choana. These fishes used their fins as paddles in shallow-water habitats choked with plants and detritus. Their fins could also have been used to attach themselves to plants or similar while they were laying in ambush for prey. The universal tetrapod characteristics of front limbs that bend backward at the elbow and hind limbs that bend forward at the knee can plausibly be traced to early tetrapods living in shallow water.
It is now clear that the common ancestor of the bony fishes had a primitive air-breathing lung (later evolved into a swim bladder in most ray-finned fishes). This suggests that it evolved in warm shallow waters, the kind of habitat the lobe finned fishes were living and made use of their simple lung when the oxygen level in the water became too low.
The lungfishes are now considered as being the closest living relatives of the tetrapods, even closer than the coelacanth.
Fleshy lobe fins supported on bones rather than ray-stiffened fins seems to have been an original trait of the bony fishes (Osteichthyes). The lobe-finned ancestors of the tetrapods evolved them further, while the ancestors of the ray-finned (Actinopterygii) fishes evolved their fins in the opposite direction. The most primitive group of the ray-fins, the bichirs, still have fleshy frontal fins.
Nine genera of Devonian tetrapods have been described, several known mainly or entirely from lower jaw material. All of them were from the European-North American supercontinent, which comprised Europe, North America and Greenland. The only exception is a single Gondwanan genus, Metaxygnathus, which has been found in Australia.
The first Devonian tetrapod identified from Asia was recognized from a fossil jawbone reported in 2002. The Chinese tetrapod Sinostega pani was discovered among fossilized tropical plants and lobe-finned fish in the red sandstone sediments of the Ningxia Hui Autonomous Region of northwest China. This finding substantially extended the geographical range of these animals and has raised new questions about the worldwide distribution and great taxonomic diversity they achieved within a relatively short time.
These earliest tetrapods were not terrestrial. The earliest confirmed terrestrial forms are known from the early Carboniferous deposits, some 20 million years later. Still, they may have spent very brief periods out of water and would have used their legs to paw their way through the mud.
Why they went to land in the first place is still debated. One reason could be that the small juveniles who had completed their metamorphosis had what it took to make use of what land had to offer. Already adapted to breathe air and move around in shallow waters near land as a protection (just as modern fish (and amphibians) often spent the first part of their life in the comparative safety of shallow waters like mangrove forests), two very different niches partially overlapped each other, with the young juveniles in the diffuse line between. One of them was overcrowded and dangerous while the other was much safer and much less crowded, offering less competition over resources. The terrestrial niche was also a much more challenging place for primary aquatic animals, but because of the way evolution and the selection pressure works, those juveniles who could take advantage of this would be rewarded. Once they gained a small foothold on land, evolution took care of the rest, thanks to all their preadaptations and being at the right place at the right time.


{kind=link}
The basal tetrapod Ichthyostega, from the Famennian (latest Devonian) of Greenland.
At this time there were a lot of invertebrates crawling around on land and near water, in moist soil and wet litter, more than big enough to give the small ones a good meal. Some were even big enough to eat small tetrapods, but land would still be a much safer place and offer more than the waters if they knew how to make use of it.

Tulerpeton curtum, from the Late Devonian of Tula Region of Russia.
Adults would be too heavy and slow and demand bigger prey. Small juveniles were much lighter, faster and was satisfied with relatively small invertebrates. Modern mudskippers are said to be able to snap insects in flight while on land, so maybe we shouldn't underestimate the early juvenile tetrapods either.
Initially making only tentative forays onto land, as the generations went by they adapted to terrestrial environments and spent longer periods away from the water, also spending a longer part of their childhood on land before returning to the water for the rest of their life. It is possible also the adults started to spend some time on land as the skeletal modifications in early tetrapods as Ichthyostega suggests, but only to bask in the sun close to the water's edge, not to hunt or move around. It is a fact that the first true tetrapods adapted to terrestrial locomotion were small. Only later did they increase in size.
The fully grown obviously kept most of the anatomical and other forms of adaptations from their juvenile stage, giving them modified limbs and other traits of terrestrial properties. To be successful adults they first had to be successful juveniles. The adults of some of the smaller species were in that case probably able to move on land too when sufficiently evolved.
If some sort of neoteny or dwarfism occurred, making the animals sexually mature and fully grown while still living on land, they would only need to visit water to drink and reproduce.
Carboniferous tetrapods[]


{kind=link}
The large Early Carboniferous basal tetrapod Crassigyrinus scoticus. This large (2 meeters long) aquatic animal lived an eel- or pike- like existence in Carboniferous swamps.
Until the 1990s, there was a 30-million year gap in the fossil record between the late Devonian tetrapods and the reappearance of tetrapod fossils in recognizable mid-Carboniferous amphibian lineages. It was referred to as "Romer's Gap", after the palaeontologist who recognized it.
During the "gap", tetrapod backbones developed, as did limbs with digits and other adaptations for terrestrial life. Ears, skulls and vertebral columns all underwent changes too. The number of digits on hands and feet became standardized at five, as lineages with more digits died out. The very few tetrapod fossils found in the "gap" are all the more precious.
The transition from an aquatic lobe-finned fish to an air-breathing amphibian was a momentous occasion in the evolutionary history of the vertebrates. For an animal to live in a gravity-neutral, aqueous environment and then invade one that is entirely different required major changes to the overall body plan, both in form and in function. Eryops is an example of an animal that made such adaptations. It retained and refined most of the traits found in its fish ancestors. Sturdy limbs supported and transported its body while out of water. A thicker, stronger backbone prevented its body from sagging under its own weight. Also, by utilizing vestigial fish jaw bones, a rudimentary ear was developed, allowing Eryops to hear airborne sound.
By the Visean age of mid-Carboniferous times the early tetrapods had radiated into at least three main branches. Recognizable basal-group Amphibia are representative of the labyrinthodonts, which are comprised of the temnospondyls (e.g. Eryops) and similarly primitive anthracosaurs, who were the relatives and ancestors of the Amniota. Depending on whichever authorities one follows, modern amphibians (frogs, salamanders and caecilians) are derived from one or the other (or possibly both, although this is now a minority position) of these two groups. The first amniotes are known from the early part of the Late Carboniferous, and during the Triassic countered among their number the earliest mammals, turtles, and crocodiles (lizards and birds appeared in the Jurassic, and snakes in the Cretaceous). As living members of the tetrapod clan—that is of the tetrapod "crown-group"—these varied tetrapods represent the phylogenetic end-points of these two divergent lineages. A third, more primitive, Carboniferous group, the baphetids, left no modern survivors. Finally, the Lepospondyli are an extinct Palaeozoic group of uncertain relationships.
Permian tetrapods[]
In the Permian period, as the separate tetrapod lineages each developed in their own way, the term "tetrapoda" becomes less useful. In addition to temnospondyl and anthracosaur clades among the early "amphibia" (labyrinthodonts), there were two important divergent clades of amniotes, the Sauropsida and the Synapsida, of which the latter were the most important and successful Permian animals. Each of these lineages, however, remains grouped with the tetrapoda, just as Homo sapiens could be considered a very highly-specialized kind of lobe-finned fish.
Mesozoic, Cenozoic and Extant tetrapods[]
There are three main categories of living ("crown group") tetrapods:
- Lissamphibia
- frogs and toads, newts and salamanders
- Sauropsida
- dinosaurs, most modern reptiles, turtles, and birds
- Synapsida
- pelycosauria, therapsida, and mammals
Note that snakes and other legless reptiles are considered tetrapods because they are descended from ancestors who had a full complement of limbs. Similar considerations apply to aquatic mammals.
Most tetrapods today are terrestrial, at least in their adult forms, but some species, such as the axolotl, remain aquatic. Tetrapods that returned to the sea include ichthyosaurs and modern whales and dolphins.
Tetrapod physiology[]
- from Wikipedia:
Anatomical features of early tetrapods[]
The amphibian's ancestral fish must have possessed similar traits to those inherited by the early amphibians, including internal nostrils (to separate the breathing and feeding passages) and a large fleshy fin built on bones that could give rise to the tetrapod limb. The rhipidistian crossopterygians fulfill every requirement for this ancestry. Their palatal and jaw structures were identical to those of amphibians, and their dentition was identical too, with labyrinthine teeth fitting in a pit-and-tooth arrangement on the palate. The crossopterygian paired fins were smaller than tetrapod limbs, but the skeletal structure was very similar in that the crossopterygian had a single proximal bone (analogous to the humerus or femur), two bones in the next segment (forearm or lower leg), and an irregular subdivision of the fin, roughly comparable to the structure of the carpus / tarsus and phalanges of a hand.
The major difference between crossopterygians and amphibians was in relative development of front and back skull portions; the snout is much less developed than in most amphibians and the post-orbital skull is exceptionally longer than an amphibian's.
A great many kinds of early amphibians lived during the Carboniferous period. Therefore, their ancestor would have lived earlier, during the Devonian period. Devonian Ichthyostegids were the earliest of amphibians, with a skeleton that is directly comparable to that of rhipidistian ancestors. Early Labyrinthodonts (Late Devonian to Early Mississippian) still had some ichthyostegid features such as similar skull bone patterns, labyrinthine tooth structure, the fish skull-hinge, pieces of gill structure between the cheek and shoulder, and the vertebral column. They had, however, lost several other fish features such as the fin rays in the tail.
In order to propagate in the terrestrial environment, certain challenges had to be overcome. The animal's body needed additional support, because buoyancy was no longer a factor. A new method of respiration was required in order to extract atmospheric oxygen, instead of oxygen dissolved in water. A means of locomotion would need to be developed to traverse distances between waterholes. Water retention was now important since it was no longer the living matrix, and it could be lost easily to the environment. Finally, new sensory input systems were required if the animal was to have any ability to function reasonably while on land.
Living tetrapods[]
The end of the Paleozoic saw a major turnover in fauna. Many of the once large and important groups died out or were greatly reduced. A group of small, generalized reptiles, the diapsids became important, notably the dinosaurs. During the Mesozoic, the old labyrinthodont amphibians died out, and the synapsids (mammal-like reptiles) shrunk to the edge of extinction and ultimately gave rise to mammals before dying out. All current major groups of reptiles evolved, and birds appeared from dinosaurs in the Jurassic.
Following the great faunal turnover at the end of the Mesozoic left only three categories of living ("crown groups") tetrapods, all of which also include many extinct groups:
- Lissamphibia: Modern frogs and toads, newts and salamanders, and caecilians
- Sauropsida: Birds and modern reptiles
- Synapsida: Mammals