
{kind=link}
An artist's depiction of a feathered Deinonychus antirrhopus, a genus of carnivorous dromaeosaurid dinosaur that lived during the early Cretaceous Period in what is now the Western US.[Note 1]
Feathered dinosaurs is a term used to describe dinosaurs, particularly maniraptoran dromaeosaurs, that were covered in plumage; either filament-like intergumentary structures with few branches, to fully developed pennaceous feathers complete with shafts and vanes. Feathered dinosaurs first came to realization after it was discovered that dinosaurs are closely related to birds. Since then, the term "feathered dinosaurs" has widened to encompass the entire concept of the dinosaur–bird relationship, including the various avian characteristics some dinosaurs possess, including a pygostyle, a posteriorly oriented pelvis, elongated arms and forelimbs and clawed hand, and clavicles fused to form a furcula. A substantial amount of evidence demonstrates that birds are the descendants of theropod dinosaurs, and that birds evolved during the Jurassic from small, feathered maniraptoran theropods closely related to dromaeosaurids and troodontids (known collectively as deinonychosaurs). Less than two dozen species of dinosaurs have been discovered with direct fossil evidence of plumage since the 1990s, with most coming from Cretaceous deposits in China, most notably Liaoning Province. Together, these fossils represent an important transition between dinosaurs and birds, which allows paleontologists to piece together the origin and evolution of birds.
Despite integumentary structures being limited to non-avian dinosaurs, particularly well-documented in maniraptoriformes, fossils do suggest that a large number of theropods were feathered, and it has even been suggested that based on phylogenetic analyses, Tyrannosaurus at one stage of its life may have been covered in down-like feathers, although there is no direct fossil evidence of this. Based on what is known of the dinosaur fossil record, paleontologists generally think that most of dinosaur evolution happened at relatively large body size (a mass greater than a few kilograms), and in animals that were entirely terrestrial. Small size (<1 kg) and arboreal habits seem to have arisen fairly late during dinosaurian evolution, and only within maniraptora.
| Part of a series on | ||
| Dinosaurs and birds | ||
|---|---|---|
 | ||
| ||
| ||
|
Birds were originally linked with other dinosaurs back in the late 1800s, most famously by Thomas Huxley. This view remained fairly popular until the 1920s when Gerhard Heilmann's book The Origin of Birds was published in English. Heilmann argued that birds could not have descended from dinosaurs (predominantly because dinosaurs lacked clavicles, or so he thought), and he therefore favored the idea that birds originated from the so-called 'pseudosuchians': primitive archosaurs that were also thought ancestral to dinosaurs and crocodilians. This became the mainstream view until the 1970s, when a new look at the anatomical evidence (combined with new data from maniraptoran theropods) led John Ostrom to successfully resurrect the dinosaur hypothesis. Fossils of Archaeopteryx include well-preserved feathers, but it was not until the early 1990s that clearly nonavian dinosaur fossils were discovered with preserved feathers. Today there are more than twenty genera of dinosaurs with fossil feathers, nearly all of which are theropods. Most are from the Yixian Formation in China. The fossil feathers of one specimen, Shuvuuia deserti, have even tested positive for beta-keratin, the main protein in bird feathers, in immunological tests.
Research history[]
Early hypotheses[]

{kind=link}
The Berlin Archaeopteryx
Shortly after the 1859 publication of Charles Darwin's The Origin of Species, the ground-breaking book which described his theory of evolution by natural selection,[1] British biologist and evolution-defender Thomas Henry Huxley proposed that birds were descendants of dinosaurs. He compared skeletal structure of Compsognathus, a small theropod dinosaur, and the 'first bird' Archaeopteryx lithographica (both of which were found in the Upper Jurassic Bavarian limestone of Solnhofen). He showed that, apart from its hands and feathers, Archaeopteryx was quite similar to Compsognathus.[2] In 1868 he published On the Animals which are most nearly intermediate between Birds and Reptiles, making the case.[3][4] The leading dinosaur expert of the time, Richard Owen, disagreed, claiming Archaeopteryx as the first bird outside dinosaur lineage.[5] For the next century, claims that birds were dinosaur descendants faded, while more popular bird-ancestry hypotheses including that of a possible 'crocodylomorph' and 'thecodont' ancestor gained ground.[6]
Aboreal dinosaurs[]


{kind=link}
Gerhard Heilmann's 'fighting proaves' reconstruction. Note the trunk-climbing 'proaves' in the background.
Since the discovery of such theropods as Microraptor and Epidendrosaurus, paleontologists and scientists in general now have small forms exhibiting some features suggestive of a tree-climbing (or scansorial) way of life.[7][8] However, the idea that dinosaurs might have climbed trees goes back a long way, and well pre-dates the dinosaur renaissance of the 1960s and 70s.
The idea of scansoriality in non-avian dinosaurs has been considered a 'fringe' idea, and it's partly for this reason that, prior to 2000, nobody had attempted any sort of review on the thoughts that had been published about the subject. The oldest reference to scansoriality in a dinosaur comes from William Fox, the Isle of Wight curator and amateur fossil collector, who in 1866 proposed that Calamospondylus oweni from the Lower Cretaceous Wessex Formation of the Isle of Wight might have been in the habit of 'leaping from tree to tree'.[9] The Calamospondylus oweni specimen that Fox referred to was lost, and the actual nature of the fossil remains speculative, but there are various reasons for thinking that it was a theropod.[10] However, it's not entirely accurate to regard Fox's ideas about Calamospondylus as directly relevant to modern speculations about tree-climbing dinosaurs given that, if Fox imagined Calamospondylus oweni as resembling anything familiar, it was probably as a lizard-like reptile, and not as a dinosaur as they are currently understood.
During the early decades of the 20th century the idea of tree-climbing dinosaurs became reasonably popular as Othenio Abel, Gerhard Heilmann and others used comparisons with birds, tree kangaroos and monkeys to argue that the small ornithopod Hypsilophodon (also from the Wessex Formation of the Isle of Wight) was scansorial. Heilmann had come to disagree with this idea and now regarded Hypsilophodon as terrestrial. William Swinton favored the idea of a scansorial Hypsilophodon, concluding that 'it would be able to run up the stouter branches and with hands and tail keep itself balanced until the need for arboreal excursions had passed',[11] and in a 1936 review of Isle of Wight dinosaurs mentioned the idea that small theropods might also have used their clawed hands to hold branches when climbing.[12]
During the 1970s, Peter Galton was able to show that all of the claims made about the forelimb and hindlimb anatomy of Hypsilophodon supposedly favoring a scansorial lifestyle were erroneous, and that this animal was in fact well suited for an entirely terrestrial, cursorial lifestyle.[13][14] Nevertheless, for several decades Hypsilophodon was consistently depicted as a tree-climber.
In recent decades, Gregory Paul has been influential in arguing that small theropods were capable climbers, and he not only argued for and illustrated scansorial abilities in coelurosaurs,[15] he also proposed that as-yet-undiscovered maniraptorans were highly proficient climbers and included the ancestors of birds. The hypothesized existence of small arboreal theropods that are as yet unknown from the fossil record later proved integral to George Olshevsky's 'Birds Came First' (BCF) hypothesis. Olshevsky argued that all dinosaurs, and in fact all archosaurs, descend from small, scansorial ancestors, and that it is these little climbing reptiles which are the direct ancestors of birds.[16][17]
Ostrom, Deinonychus and the Dinosaur Renaissance[]
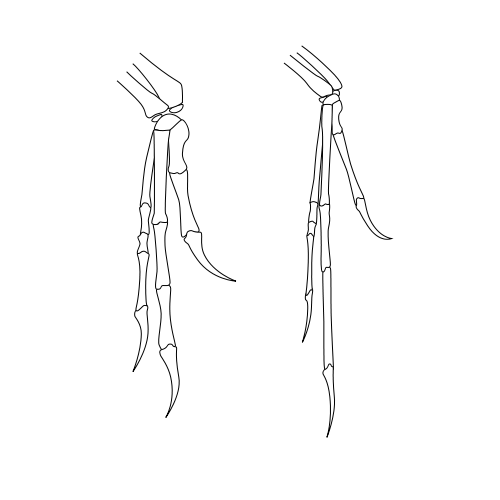

{kind=link}
The similarity of the forelimbs of Deinonychus (left) and Archaeopteryx (right) led John Ostrom to revive the link between dinosaurs and birds.
In 1964, the first specimen of Deinonychus antirrhopus[5] was discovered in Montana, and in 1969, John Ostrom of Yale University described Deinonychus as a theropod whose skeletal resemblance to birds seemed unmistakable.[18] Since that time, Ostrom had become a leading proponent of the theory that birds are direct descendants of dinosaurs. During the late 1960s, Ostrom and others demonstrated that maniraptoran dinosaurs could fold their arms in a manner similar to that of birds.[18] Further comparisons of bird and dinosaur skeletons, as well as cladistic analysis strengthened the case for the link, particularly for a branch of theropods called maniraptors. Skeletal similarities include the neck, the pubis, the wrists (semi-lunate carpal), the 'arms' and pectoral girdle, the shoulder blade, the clavicle and the breast bone. In all, over a hundred distinct anatomical features are shared by birds and theropod dinosaurs.
Other researchers drew on these shared features and other aspects of dinosaur biology and began to suggest that at least some theropod dinosaurs were feathered. The first restoration of a feathered dinosaur was Sarah Landry's depiction of a feathered "Syntarsus" (now renamed Megapnosaurus or considered a synonym of Coelophysis), in Robert T. Bakker's 1975 publication Dinosaur Renaissance.[19] Gregory S. Paul was probably the first paleoartist to depict maniraptoran dinosaurs with feathers and protofeathers, starting in the late 1980s.
By the 1990s, most paleontologists considered birds to be surviving dinosaurs and referred to 'non-avian dinosaurs' (all extinct), to distinguish them from birds (aves). Before the discovery of feathered dinosaurs, the evidence was limited to Huxley and Ostrom's comparative anatomy. Some mainstream ornithologists, including Smithsonian Institution curator Storrs L. Olson, disputed the links, specifically citing the lack of fossil evidence for feathered dinosaurs.[20]
Modern research and feathered dinosaurs in China[]
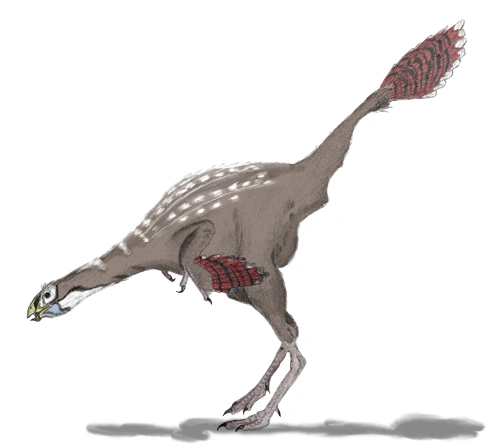

{kind=link}
Life restoration of the feathered dinosaur Caudipteryx zoui.
The early 1990s saw the discovery of spectacularly preserved bird fossils in several Early Cretaceous[21][22] geological formations in the northeastern Chinese province of Liaoning.[23][24] South American paleontologists, including Fernando Novas and others, discovered evidence showing that maniraptorans could move their arms in a bird-like manner.[25][26] Gatesy and others suggested that anatomical changes to the vertebral column and hindlimbs occured before birds first evolved,[27][28] and Xu Xing and colleagues proved that true functional wings and flight feathers evolved in some maniraptorans,[29][7] all strongly suggesting that these anatomical features were already well-developed before the first birds evolved.[30]
In 1996, Chinese paleontologists described Sinosauropteryx as a new genus of bird from the Yixian Formation,[31] but this animal was quickly recognized as a theropod dinosaur closely related to Compsognathus. Surprisingly, its body was covered by long filamentous structures. These were dubbed 'protofeathers' and considered to be homologous with the more advanced feathers of birds,[32] although some scientists disagree with this assessment.[33] Chinese and North American scientists described Caudipteryx and Protarchaeopteryx soon after. Based on skeletal features, these animals were non-avian dinosaurs, but their remains bore fully-formed feathers closely resembling those of birds.[34] "Archaeoraptor," described without peer review in a 1999 issue of National Geographic,[35] turned out to be a smuggled forgery,[36] but legitimate remains continue to pour out of the Yixian, both legally and illegally. Many newly described feathered dinosaurs preserve horny claw sheaths, integumentary structures (filaments to fully pennaceous feathers), and internal organs.[30] Feathers or "protofeathers" have been found on a wide variety of theropods in the Yixian,[37][38] and the discoveries of extremely bird-like dinosaurs,[7] as well as dinosaur-like primitive birds,[39] have almost entirely closed the morphological gap between theropods and birds.
Features linking birds and dinosaurs[]
Over one hundred distinct anatomical features are shared by birds and theropod dinosaurs.[Note 2] Some of the more interesting and notable similarities are discussed here:
Feathers[]
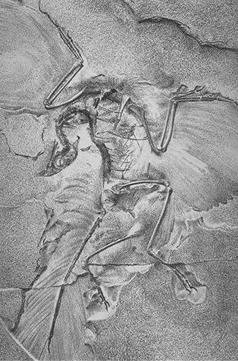

{kind=link}
The Berlin Archaeopteryx, 1881.
Archaeopteryx, the first good example of a "feathered dinosaur", was discovered in 1861. The initial specimen was found in the solnhofen limestone in southern Germany, which is a lagerstätte, a rare and remarkable geological formation known for its superbly detailed fossils. Archaeopteryx is a transitional fossil, with features clearly intermediate between those of modern reptiles and birds. Discovered just two years after Darwin's seminal Origin of Species, its discovery spurred the nascent debate between proponents of evolutionary biology and creationism. This early bird is so dinosaur-like that, without a clear impression of feathers in the surrounding rock, at least one specimen was mistaken for Compsognathus.[40]
Since the 1990s, a number of additional feathered dinosaurs have been found, providing even stronger evidence of the close relationship between dinosaurs and modern birds. Most of these specimens were unearthed in Liaoning province, northeastern China, which was part of an island continent during the Cretaceous period.[41] Though feathers have been found only in the lagerstätte of the Yixian Formation and a few other places, it is possible that non-avian dinosaurs elsewhere in the world were also feathered. The lack of widespread fossil evidence for feathered non-avian dinosaurs may be due to the fact that delicate features like skin and feathers are not often preserved by fossilization and thus are absent from the fossil record.


{kind=link}
Fossil cast of NGMC 91, a probable specimen of Sinornithosaurus.
A recent development in the debate centers around the discovery of impressions of "protofeathers" surrounding many dinosaur fossils. These protofeathers suggest that the tyrannosauroids may have been feathered.[38] However, others claim that these protofeathers are simply the result of the decomposition of collagenous fiber that underlaid the dinosaurs' integument.[42] The Dromaeosauridae family, in particular, seems to have been heavily feathered and at least one dromaeosaurid, Cryptovolans, may have been capable of flight.[43][44]
Skeleton[]
Because feathers are often associated with birds, feathered dinosaurs are often touted as the missing link between birds and dinosaurs. However, the multiple skeletal features also shared by the two groups represent the more important link for paleontologists. Furthermore, it is increasingly clear that the relationship between birds and dinosaurs, and the evolution of flight, are more complex topics than previously realized. For example, while it was once believed that birds evolved from dinosaurs in one linear progression, some scientists, most notably Gregory S. Paul, conclude that dinosaurs such as the dromaeosaurs may have evolved from birds, losing the power of flight while keeping their feathers in a manner similar to the modern ostrich and other ratites.[45]
Comparisons of bird and dinosaur skeletons, as well as cladistic analysis, strengthens the case for the link, particularly for a branch of theropods called maniraptors. Skeletal similarities include the neck, pubis, wrist (semi-lunate carpal), arm and pectoral girdle, shoulder blade, clavicle, and breast bone.
Furcula[]


{kind=link}
Dave's wishbone is highlighted in this image.


{kind=link}
Bronze cast of the wishbone of "Sue", Field Museum.
At one time, it was believed that dinosaurs lacked furculae, long believed to be a structure unique to birds, that were formed by the fusion of the two collarbones (clavicles) into a single V-shaped structure that helps brace the skeleton against the stresses incurred while flapping. This apparent absence was considered an overwhelming argument to refute the dinosaur ancestry of birds by Danish artist and naturalist Gerhard Heilmann's monumentally influential The Origin of Birds in 1926.[46] That reptiles ancestral to birds, therefore, should, at the very least, show well-developed clavicles. In the book, Heilmann discussed that no clavicles had been reported in any theropod dinosaur. Noting this fact, Heilmann suggested that birds evolved from a more generalized archosaurian ancestor, such as the aptly-named Ornithosuchus (literally, “bird-crocodile”), which is now believed to be closer to the crocodile end of the archosaur lineage. At the time, however, Ornithosuchus seemed to be a likely ancestor of more birdlike creatures.[47]
Contrary to what Heilman believed, paleontologists since the 1980s now accept that clavicles and in most cases furculae are a standard feature not just of theropods but of saurischian dinosaurs.[48] Furculae in dinosaurs is not only limited to maniraptorans, as evidenced by an article by Chure & Madson in which they described a furcula in an allosaurid dinosaur, a non-avian theropod.[49][50][51] In 1983, Rinchen Barsbold reported the first dinosaurian furcula from a specimen of the Cretaceous theropod Oviraptor.[52] A furcula-bearing Oviraptor specimen had previously been known since the 1920s, but because at the time the theropod origin of birds was largely dismissed, it was misidentified for sixty years.[45]:9
Following this discovery, paleontologists began to find furculae in other theropod dinosaurs.[53] Wishbones are now known from the dromaeosaur Velociraptor,[6] the allosauroid Allosaurus,[7] and the tyrannosaurid Tyrannosaurus rex, to name a few. Up to late 2007, ossified furculae (i.e. made of bone rather than cartilage) have been found in nearly all types of theropods except the most basal ones, Eoraptor and Herrerasaurus.[54] The original report of a furcula in the primitive theropod Segisaurus (1936) has been confirmed by a re-examination in 2005.[55] Joined, furcula-like clavicles have also been found in Massospondylus, an Early Jurassic sauropodomorph, indicating that the evolution of the furcula was well underway when the earliest dinosaurs were diversifying.[56]
Coelophysis[]


{kind=link}
The wishbone of Coelophysis from Coelophysis Quarry at Ghost Ranch, New Mexico, 210 Ma.
In 2000, Alex Downs reported an isolated furcula found within a block of Coelophysis bauri skeletons from the Late Triassic Rock Point Formation at Ghost Ranch, New Mexico.[57] While it seemed likely that it originally belonged to Coelophysis, the block contained fossils from other Triassic animals as well, and Alex declined to make a positive identification. Currently, a total of five C. bauri furculae have been found in the New Mexico Museum of Natural History's (NMMNH) Ghost Ranch, New Mexico Whitaker Quarry block C-8-82. Three of the furculae are articulated in juvenile skeletons; two of these are missing fragments but are nearly complete, and one is apparently complete.[58][59][60] Two years later, Tykoski et al. described several furculae from two species of the coelophysoid genus Syntarsus (now Megapnosaurus), S. rhodesiensis and S. kayentakatae, from the Early Jurassic of Zimbabwe and Arizona, respectively.[59] Syntarsus was long considered to be the genus most closely related to Coelophysis, differing only in a few anatomical details and slightly younger age, so the identification of furculae in Syntarsus made it very likely that the furcula Alex Downs noted in 2000 came from Coelophysis after all. By 2006, wishbones were definitively known from the Early Jurassic Coelophysis rhodesiensis and Coelophysis kayentakatae, and a single isolated furcula was known that might have come from the Late Triassic type species, Coelophysis bauri.
Avian air sacs[]
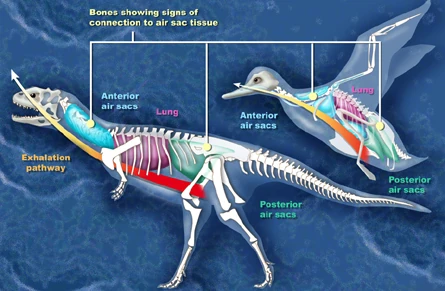
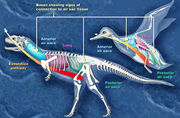
{kind=link}
A recent analysis showed the presence of a very bird-like lung system in predatory dinosaurs.[61]
Large meat-eating dinosaurs had a complex system of air sacs similar to those found in modern birds, according to an investigation which was led by Patrick O'Connor of Ohio University. The lungs of theropod dinosaurs (carnivores that walked on two legs and had birdlike feet) likely pumped air into hollow sacs in their skeletons, as is the case in birds. "What was once formally considered unique to birds was present in some form in the ancestors of birds", O'Connor said.[62] In a paper published in the online journal Public Library of Science ONE (September 29, 2008), scientists described Aerosteon riocoloradensis, the skeleton of which supplies the strongest evidence to date of a dinosaur with a bird-like breathing system. CT-scanning revealed the evidence of air sacs within the body cavity of the Aerosteon skeleton.[61][63]
Heart and sleeping posture[]


{kind=link}
Mei slept with its snout nestled beneath one of the forelimbs, similar to modern birds.
Modern computed tomography (CT) scans of a dinosaur chest cavity conducted in 2000 found the apparent remnants of complex four-chambered hearts, much like those found in today's mammals and birds.[64] The idea is controversial within the scientific community, coming under fire for bad anatomical science[65] or simply wishful thinking.[66] The type fossil of the troodont, Mei, is complete and exceptionally well preserved in three-dimensional detail, with the snout nestled beneath one of the forelimbs, similar to the roosting position of modern birds. This demonstrates that the dinosaurs slept like certain modern birds, with their heads tucked under their arms.[67] This behavior, which may have helped to keep the head warm, is also characteristic of modern birds.
Reproductive biology[]
A discovery of features in a Tyrannosaurus rex skeleton recently provided more evidence that dinosaurs and birds evolved from a common ancestor and, for the first time, allowed paleontologists to establish the sex of a dinosaur. When laying eggs, female birds grow a special type of bone in their limbs between the hard outer bone and the marrow. This medullary bone, which is rich in calcium, is used to make eggshells. The presence of endosteally derived bone tissues lining the interior marrow cavities of portions of the Tyrannosaurus rex specimen's hind limb suggested that T. rex used similar reproductive strategies, and revealed the specimen to be female.[68] Further research has found medullary bone in the theropod Allosaurus and ornithopod Tenontosaurus. Because the line of dinosaurs that includes Allosaurus and Tyrannosaurus diverged from the line that led to Tenontosaurus very early in the evolution of dinosaurs, this suggests that dinosaurs in general produced medullary tissue. Medullary bone has been found in specimens of sub-adult size, which suggests that dinosaurs reached sexual maturity rather quickly for such large animals.[69] The micro-structure of eggshells and bones has also been determined to be similar to that of birds.[70]
Brooding and care of young[]


{kind=link}
A nesting Citipati osmolskae specimen, at the American Museum of Natural History in New York.
Several specimens of the Mongolian oviraptorid Citipati was discovered in a chicken-like brooding position resting over the eggs in its nest in 1993, which may mean it was covered with an insulating layer of feathers that kept the eggs warm.[71][72][73][74] All of the nesting specimens are situated on top of egg clutches, with their limbs spread symmetrically on each side of the nest, front limbs covering the nest perimeter. This brooding posture is found today only in birds and supports a behavioral link between birds and theropod dinosaurs.[72] The nesting position of Citipati also supports the hypothesis that it and other oviraptorids had feathered forelimbs. With the 'arms' spread along the periphery of the nest, a majority of eggs would not be covered by the animal's body unless an extensive coat of feathers was present.[45]
A dinosaur embryo was found without teeth, which suggests some parental care was required to feed the young dinosaur, possibly the adult dinosaur regurgitated food into the young dinosaur's mouth (see altricial).[75] This behavior is seen in numerous bird species; parent birds regurgitate food into the hatchling's mouth.
Teeth[]
The loss of teeth and the formation of a beak has been shown to have been favorably selected to suit the newly aerodynamical bodies of avian flight in early birds.[76] In the Jehol Biota in China, various dinosaur fossils have been discovered that have a variety of different tooth morphologies, in respect to this evolutionary trend. Sinosauropteryx fossils display unserrated premaxillary teeth, while the maxillary teeth are serrated.[32] In the preserved remains of Protarchaeopteryx, four premaxillary teeth are present that are serrated.[77] The diminutive oviraptorosaur Caudipteryx has four hook-like premaxillary teeth,[34] and in Microraptor zhaoianus, the posterior teeth of this species had developed a constriction that led to a less compressed tooth crown.[78] These dinosaurs exhinit a heterodont dentition pattern that clearly illustrates a transition from the teeth of maniraptorans to those of early, basal birds.[78]
Molecular evidence and soft tissue[]


{kind=link}
Fossil of a juvenile individual of Scipionyx samniticus. The fossil preserves clear traces of soft tissues.
One of the best examples of soft tissue impressions in a fossil dinosaur was discovered in Petraroia, Italy. The discovery was reported in 1998, and described the specimen of a small, very young coelurosaur, Scipionyx samniticus. The fossil includes portions of the intestines, colon, liver, muscles, and windpipe of this immature dinosaur.[79]
In the March 2005 issue of Science, Dr. Mary Higby Schweitzer and her team announced the discovery of flexible material resembling actual soft tissue inside a 68-million-year-old Tyrannosaurus rex leg bone from the Hell Creek Formation in Montana. After recovery, the tissue was rehydrated by the science team. The seven collagen types obtained from the bone fragments, compared to collagen data from living birds (specifically, a chicken), reveal that older theropods and birds are closely related.[80]
When the fossilized bone was treated over several weeks to remove mineral content from the fossilized bone marrow cavity (a process called demineralization), Schweitzer found evidence of intact structures such as blood vessels, bone matrix, and connective tissue (bone fibers). Scrutiny under the microscope further revealed that the putative dinosaur soft tissue had retained fine structures (microstructures) even at the cellular level. The exact nature and composition of this material, and the implications of Dr. Schweitzer's discovery, are not yet clear; study and interpretation of the specimens is ongoing.[81]
The successful extraction of ancient DNA from dinosaur fossils has been reported on two separate occasions, but upon further inspection and peer review, neither of these reports could be confirmed.[82] However, a functional visual peptide of a theoretical dinosaur has been inferred using analytical phylogenetic reconstruction methods on gene sequences of related modern species such as reptiles and birds.[83] In addition, several proteins have putatively been detected in dinosaur fossils,[84] including hemoglobin.[85]
Evolution[]
Feathers are extremely complex integumentary structures that characterize a handful of vertebrate animals.[30][86][87] Although it is generally acknowledged that feathers are derived and evolved from simpler integumentary structures, the early diversification and origin of feathers was relatively unknown until recently, and current research is ongoing. Since the theropod ancestry of birds is widely supported with osteological and other physical lines of evidence, the precursors of feathers in dinosaurs are also present, as predicted by those who originally proposed a theropod origin for birds.[88] In 2006, Chinese paleontologist Xu Xing stated in a paper that since many members of Coelurosauria exhibit miniaturization, primitive integumentary structures (and later on feathers) evolved in order to insulate their small bodies.[30]
Original function[]
The functional view on the evolution of feathers has traditionally focussed on insulation, flight and display. Discoveries of non-flying Late Cretaceous feathered dinosaurs in China however suggest that flight could not have been the original primary function.[89] Feathers in dinosaurs indicate that their original function was not flight, but of a different nature. Theories include insulation brought around after they had metabolically changed from their cold-blooded reptilian ancestors, to increasing running speed.[90] It has been suggested that vaned feathers evolved in the context of thrust, with running, non-avian theropods flapping their arms to increase their running speed.[91]
Evolutionary models[]

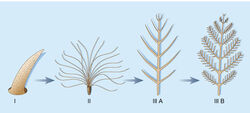
{kind=link}
Hypothesized stages I–III of feather evolution.
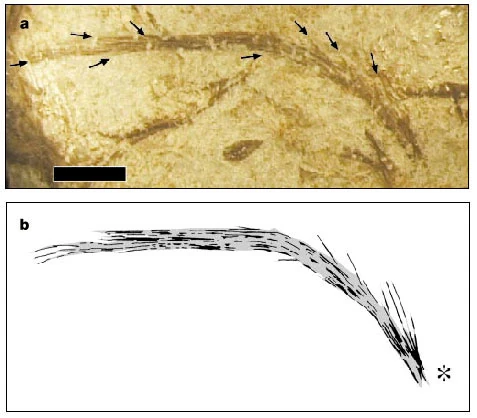
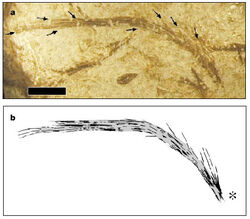
{kind=link}
Filamentous integumental structure of Sinornithosaurus millenii with compound structures composed of multiple filaments may represent Stage II in the evolution of feathers.
The following is the generally acknowledged version of the origin and early evolution of feathers:[30]
- The first feathers evolved; they are single filaments.
- Branching structures developed.
- The rachis evolved.
- Pennaceous feathers evolved.
- Aerodynamic morphologies appeared. (curved shaft and asymmetrical vanes)
This scenario appears to indicate that downy, contour, and flight feathers, are more derived forms of the first "feather".[30] However, it is also possible that protofeathers and basal feathers disappeared early on in the evolution of feathers and that more primitive feathers in modern birds are secondary. This would imply that the feathers in modern birds have nothing to do with protofeathers.[30]
A recent study performed by Prum and Brush (2002) suggested that the feathers of birds are not homologous with the scales of reptiles.[92] A new model of feather evolution posits that the evolution of feathers first began with a feather follicle merging from the skin's surface that has no relation to reptilian scales.[30] After this initial event, additions and new morphological characteristics were added to the feather design and more complex feathers evolved.[93][92] This model of feather evolution, while agreeing with the distribution of various feather morphologies in coelurosaurs, it is also at odds with other evidence. The feather bristles of modern-day turkeys resemble the hair-like integumentary strcutures found in some maniraptorans, pterosaurs (see Pterosauria#Pycnofibers), and ornithischians, are widely regarded to be homologous to modern feathers, yet share also show distinct, feather like characteristics. This has led some paleontologists, such as Xu Xing, to theorize that feathers share homology with lizard scales after all.[30]
In a 2006 paper, Xu Xing discussed his idea of a new model of feather evolution:[30]
- Stage I: Tubular filaments and feather-type beta keratin evolved.[Note 3]
- Stage II: The filamentous structure evolved distal branches.[Note 4]
- Stage III: Xu Xing described this stage as being the most important stage. The main part of the modern feather, the feather follicle, appeared along with the rachises and planar forms developed.[Note 5]
- Stage IV: Large, stiff, well-developed pennaceous feathers evolved on the limbs and tails of maniraptoran dinosaurs. Barbules evolved.[Note 6]
- Stage V: Feather tracts (pennaceous feathers that are located on regions other than the limbs and tail) evolved. Specialized pennaceous feathers developed.
Xu Xing himself stated that this new model was similar to the one out forward by Richard Prum,[93][30] with the exception that Xu's model posits that feathers "feature a combination of transformation and innovation".[30] This view differs from Prum's model in that Prum suggested that feathers were purely an evolutionary novelty.[93] Xing's new model also suggests that the tubular filaments and branches evolved before the appearance of the feather follicle, while also acknowledging that the follicle was an important development in feather evolution,[30] also in contrast to Prum's model of feather evolution.
Primitive feather types[]


{kind=link}
Feather growth began with a thickening of the epidermis (skin), which elongated into a hollow tapered tube. These tubes became "protofeathers."
The evolution of feather structures is thought to have proceeded from simple hollow filaments through several stages of increasing complexity, ending with the large, deeply rooted, feathers with strong pens (rachis), barbs and barbules that birds display today.[92] It is logical that the simplest structures were probably most useful as insulation, and that this implies homeothermy. Only the more complex feather structures would be likely candidates for aerodynamic uses.
Models of feather evolution are often proposed that the earliest prototype feathers were hair-like integumentary filaments similar to the structures of Sinosauropteryx, a compsognathid (Jurassic/Cretaceous, 150-120 Ma), and Dilong, a basal tyrannosauroid from the Early Cretaceous.[93][94][92] It is not known with certainty at what point in archosaur phylogeny the earliest simple “protofeathers” arose, or if they arose once or, independently, multiple times. Filamentous structures are clearly present in pterosaurs, and long, hollow quills have been reported in a specimen of Psittacosaurus from Liaoning.[95] It is thus possible that the genes for building simple integumentary structures from beta keratin arose before the origin of dinosaurs, possibly in the last common ancestor with pterosaurs – the basal Ornithodire.
In Prum's model of feather evolution, hollow quill-like integumentary structures of this sort were termed Stage 1 feathers. The idea that feathers started out as hollow quills also supports Alan Brush's idea that feathers are evolutionary novelties, and not derived from scales. However, in order to determine the homology of Stage 1 feathers, it is necessary to determine their proteinaceous content: unlike the epidermal appendages of all other vertebrates, feathers are almost entirely composed of beta-keratins (as opposed to alpha-keratins) and, more specifically, they are formed from a group of beta-keratins called phi-keratins.[96] No studies have yet been performed on the Stage 1 structures of Sinosauropteryx or Dilong in order to test their proteinaceous composition, however, tiny filamentous structures discovered adjacent to the bones of the alvarezsaurid Shuvuuia have been tested for beta-keratin, and the structures were discovered to be composed of beta-keratin.[96][97] Alvarezsaurids have been of controversial phylogenetic position, but are generally agreed to be basal members of the Maniraptora clade. Due to this discovery, paleontologists are now convinced that beta-keratin-based protofeathers had evolved at the base of this clade at least.
Vaned, pennaceous feathers[]


{kind=link}
A reconstruction of Microraptor gui in gliding flight.
While basal coelurosaurs possessed these apparently hollow quill-like 'Stage 1' filaments, they lacked the more complex structures seen in maniraptorans. Maniraptorans possessed vaned feathers with barbs, barbules and hooklets just like those of modern birds.
The first dinosaur fossils from the Yixian formation found to have true flight-structured feathers (pennaceous feathers) were Protarchaeopteryx and Caudipteryx (135-121 Ma). Due to the size and proportions of these animals it is more likely that their feathers were used for display rather than for flight. Subsequent dinosaurs found with pennaceous feathers include Pedopenna and Jinfengopteryx. Several specimens of Microraptor, described by Xu et al. in 2003, show not only pennaceous feathers but also true asymmetrical flight feathers, present on the fore and hind limbs and tail.[7] Asymmetrical feathers are considered important for flight in birds. Before the discovery of Microraptor gui, Archaeopteryx was the most primitive known animal with asymmetrical flight feathers.[7]
However, the bodies of maniraptorans were not covered in vaned feathers as are those of the majority of living birds: instead, it seems that they were at least partly covered in the more simple structures that they had inherited from basal coelurosaurs like Sinosauropteryx. This condition may have been retained all the way up into basal birds: despite all those life restorations clothing archaeopterygids in vaned breast, belly, throat and neck feathers, it seems that their bodies also were at least partly covered in the more simple filamentous structures. The Berlin Archaeopteryx specimen appears to preserve such structures on the back of the neck though pennaceous vaned feathers were present on its back, at least.[98]
Though it has been suggested at times that vaned feathers simply must have evolved for flight, the phylogenetic distribution of these structures currently indicates that they first evolved in flightless maniraptorans and were only later exapted by long-armed maniraptorans for use in locomotion. Of course a well-known minority opinion, best known from the writings of Gregory Paul, is that feathered maniraptorans are secondarily flightless and descend from volant bird-like ancestors.[45] While this hypothesis remains possible, it lacks support from the fossil record, though that may or may not mean much, as the fossil record is incomplete and prone to selection bias.[99]
Display feathers[]


{kind=link}
Epidexipteryx represents the earliest known example of ornamental feathers in the fossil record.[100][101]
The discovery of Epidexipteryx represented the earliest known examples of ornamental feathers in the fossil record.[101] Epidexipteryx is known from a well preserved partial skeleton that includes four long feathers on the tail, composed of a central rachis and vanes. However, unlike in modern-style rectrices (tail feathers), the vanes were not branched into individual filaments but made up of a single ribbon-like sheet. Epidexipteryx also preserved a covering of simpler body feathers, composed of parallel barbs as in more primitive feathered dinosaurs. However, the body feathers of Epidexipteryx are unique in that some appear to arise from a "membranous structure."[101] The skull of Epidexipteryx is also unique in a number of features, and bears an overall similarity to the skull of Sapeornis, oviraptorosaurs and, to a lesser extent, therizinosauroids. The tail of Epidexipteryx bore unusual vertebrae towards the tip which resembled the feather-anchoring pygostyle of modern birds and some oviraptorosaurs.[101] Despite its close relationship to avialan birds, Epidexipteryx appears to have lacked remiges (wing feathers), and it likely could not fly. Zhang et al. suggest that unless Epidexipteryx evolved from flying ancestors and subsequently lost its wings, this may indicate that advanced display feathers on the tail may have predated flying or gliding flight.[101]
According to the model of feather evolution developed by Prum & Brush, feathers started out ('stage 1') as hollow cylinders, then ('stage 2') became unbranched barbs attached to a calamus. By stage 3, feathers were planar structures with the barbs diverging from a central rachis, and from there pennaceous feathers.[92][102] The feathers of Epidexipteryx may represent stage 2 structures, but also suggests that a more complicated sequence of steps in the evolution of feathers took place.[103]
Use in predation[]
Several maniraptoran lineages were clearly predatory and, given the morphology of their manual claws, fingers and wrists, presumably in the habit of grabbing at prey with their hands. Contrary to popular belief, feathers on the hands would not have greatly impeded the use of the hands in predation. Because the feathers are attached at an angle roughly perpendicular to the claws, they are oriented tangentially to the prey's body, regardless of prey size.[104]:315 It is important to note here that theropod hands appear to have been oriented such that the palms faced medially (facing inwards), and were not parallel to the ground as used to be imagined.[90]
However, feathering would have interfered with the ability of the hands to bring a grasped object up toward the mouth given that extension of the maniraptoran wrist would have caused the hand to rotate slightly upwards on its palmar side. If both feathered hands are rotated upwards and inwards at the same time, the remiges from one hand would collide with those of the other. For this reason, maniraptorans with feathered hands could grasp objects, but would probably not be able to carry them with both hands.[90] However, dromaeosaurids and other maniraptorans may have solved this problem by clutching objects single-handedly to the chest.[105] Feathered hands would also have restricted the ability of the hands to pick objects off of the ground, given that the feathers extend well beyond the ends of the digits. It remains possible that some maniraptorans lacked remiges on their fingers, but the only evidence available indicates the contrary. It has recently been argued that the particularly long second digit of the oviraptorosaur Chirostenotes was used as a probing tool, locating and extracting invertebrates and small mammals and so on from crevices and burrows.[106] It seems highly unlikely that a digit that is regularly thrust into small cavities would have had feathers extending along its length, so either Chirostenotes didn't probe as proposed, or its second finger was unfeathered, unlike that of Caudipteryx and the other feathered maniraptorans. Given the problems that the feathers might have posed for clutching and grabbing prey from the ground, we might also speculate that some of these dinosaurs deliberately removed their own remiges by biting them off.[90] Some modern birds (notably motmots) manipulate their own feathers by biting off some of the barbs, so this is at least conceivable, but no remains in the fossil record have been recovered that support this conclusion.
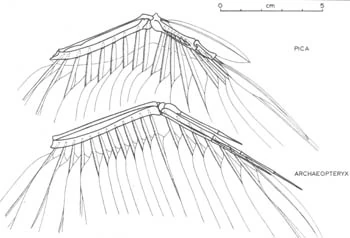
{kind=link}
Yalden's reconstruction of Archaeopteryx, with a modern magpie's wing above it.[107]
Homology and comparison to modern feathers[]
Some feather morphologies in non-avian theropods are comparable to those on modern birds. Single filament like structures are not present in modern feathers, although some birds possess highly specialized feathers that are superficial in appearance to protofeathers in non-avian theropods.[30] Tuft-like structures seen in some maniraptorans are similar to that of the natal down in modern birds.[30] Similarly, structures in the fossil record composed of a series of filaments joined at their bases along a central filament bear an uncanny resemblance to the down feathers in modern birds, with the exception of a lack of barbules. Furthermore, structures on fossils have been recovered from Chinese Cretaceous deposits that are a series of filaments joined at their bases at the distal portion of the central filament bear a superficial resemblance to filoplumes.[30] More derived, pennaceous, feathers on the tails and limbs of feathered dinosaurs are nearly identical to the remiges and retrices of modern birds.[30]
Feather structures and anatomy[]
Feathers vary in length according to their position on the body, with the filaments of the compsognathid Sinosauropteryx being 13 mm and 21 mm long on the neck and shoulders respectively. In contrast, the structures on the skull are about 5 mm long, those on the arm about 2 mm long, and those on the distal part of the tail about 4 mm long.[108][109] Because the structures tend to be clumped together it is difficult to be sure of an individual filament's morphology. The structures might have been simple and unbranched, but Currie & Chen (2001) thought that the structures on Sinosauropteryx might be branched and rather like the feathers of birds that have short quills but long barbs.[109] The similar structures of Dilong also appear to exhibit a simple branching structure.[38]
Arrangement[]
Exactly how feathers were arranged on the arms and hands of both basal birds and non-avian maniraptorans has long been unclear, and both non-avian maniraptorans and archaeopterygids have conventionally been depicted as possessing unfeathered fingers. However, the second finger is needed to support the remiges,[Note 7] and therefore must have been feathered. Derek Yalden's 1985 study was important in showing exactly how the remiges would have grown off of the first and second phalanges of the archaeopterygid second finger,[107] and this configuration has been widely recognized.[110][111][112][113][114]:129-159
However, there has been some minor historical disagreement over exactly how many remiges were present in archaeopterygids (there were most likely 11 primaries and a tiny distal 12th one, and at least 12 secondaries), and also about how the hand claws were arranged. The claws were directed perpendicularly to the palmar surface in life, and rotated anteriorly in most (but not all) specimens during burial.[114]:129-159 It has also been suggested on occasion that the fingers of archaeopterygids and other feathered maniraptorans were united in a single fleshy 'mitten' as they are in modern birds, and hence unable to be employed in grasping.[115] However, given that the interphalangeal finger joints of archaeopterygids appear suited for flexion and extension, and that the third finger apparently remained free and flexible in birds more derived than archaeopterygids,[112] this is unlikely to be correct; it's based on a depression in the sediment that was identified around the bones.
Like those of archaeopterygids and modern birds, the remiges of non-avian theropods would also have been attached to the phalanges of the second manual digit as well as to the metacarpus and ulna, and indeed we can see this in the fossils. It's the case in the sinornithosaur NGMC 91-A and Microraptor. Surprisingly, in Caudipteryx, the remiges are restricted to the hands alone, and don't extend from the arm. They seem to have formed little 'hand flags' that are unlikely to have served any function other than display.[34] Caudipteryx is an oviraptorosaur and possesses a suite of characters unique to this group. It is not a member of Aves, despite the efforts of some workers to make it into one.[116][117] The hands of Caudipteryx supported symmetrical, pennaceous feathers that had vanes and barbs, and that measured between 15–20 centimeters long (6–8 inches). These primary feathers were arranged in a wing-like fan along the second finger, just like primary feathers of birds and other maniraptorans. No fossil of Caudipteryx preserves any secondary feathers attached to the forearms, as found in dromaeosaurids, Archaeopteryx and modern birds.[118] Either these arm feathers are not preserved, or they were not present on Caudipteryx in life. An additional fan of feathers existed on its short tail. The shortness and symmetry of the feathers, and the shortness of the arms relative to the body size, indicate that Caudipteryx could not fly. The body was also covered in a coat of short, simple, down-like feathers.
Skepticism[]
A small minority, including ornithologists Alan Feduccia and Larry Martin, continues to assert that birds are instead the descendants of earlier archosaurs, such as Longisquama or Euparkeria.[119][120][42] Embryological studies of bird developmental biology have raised questions about digit homology in bird and dinosaur forelimbs.[121]
Opponents also claim that the dinosaur-bird hypothesis is dogma, apparently on the grounds that those who accept it have not accepted the opponents' arguments for rejecting it.[122] However, science does not require unanimity and does not force agreement, nor does science settle issues by vote. It has been over 25 years since John Ostrom first put forth the dinosaur-bird hypothesis in a short article in Nature,[123] and the opponents of this theory have yet to propose an alternative, testable hypothesis.[122][6] However, due to the cogent evidence provided by comparative anatomy and phylogenetics, as well as the dramatic feathered dinosaur fossils from China, the idea that birds are derived dinosaurs, first championed by Huxley and later by Nopcsa and Ostrom, enjoys near-unanimous support among today's paleontologists.[124]
BADD, BAND, and the Birds Came First hypothesis[]
- Main article: Birds Came First


{kind=link}
The "Birds Came First" hypothesis of bird and dinosaur evolution.
The non-standard, non-mainstream Birds Came First (or BCF) hypothesis proposed by George Olshevsky suggests that while there is a close relationship between dinosaurs and birds, but argues that, merely given this relationship, it is just as likely that dinosaurs descended from birds as the other way around.[125] The hypothesis does not propose that birds in the proper sense evolved earlier than did other dinosaurs or other archosaurs: rather, it posits that small, bird-like, arboreal archosaurs were the direct ancestors of all the archosaurs that came later on (proper birds included).[16][17] George was aware of this fact, and apparently considered the rather tongue-in-cheek alternative acronym GOODD, meaning George Olshevsky On Dinosaur Descendants. This was, of course, meant as opposite to the also tongue-in-cheek BADD (Birds Are Dinosaur Descendants): the term George uses for the 'conventional' or 'mainstream' view of avian origins outlined in the first two paragraphs above. 'BADD' is bad, according to BCF, as it imagines that small size, feathers and arboreal habits all evolved very late in archosaur evolution, and exclusively within maniraptoran theropod dinosaurs.[126]
Protoavis[]


{kind=link}
Protoavis restoration from Chatterjee (1999).[127]
Protoavis is a Late Triassic archosaurian whose fossilized remains were found near Post, Texas. These fossils have been described as a primitive bird which, if the identification is valid, would push back avian origins some 60-75 million years.[128][129][127]
Though it existed far earlier than Archaeopteryx, its skeletal structure is allegedly more bird-like.[130] The fossil bones are too badly preserved to allow an estimate of flying ability; although reconstructions usually show feathers, judging from thorough study of the fossil material there is no indication that these were present.[131][132]
However, this description of Protoavis assumes that Protoavis has been correctly interpreted as a bird. Almost all paleontologists doubt that Protoavis is a bird, or that all remains assigned to it even come from a single species, because of the circumstances of its discovery and unconvincing avian synapomorphies in its fragmentary material.[133][134][135][136] When they were found at a Dockum Formation quarry in the Texas panhandle in 1984, in a sedimentary strata of a Triassic river delta, the fossils were a jumbled cache of disarticulated bones that may reflect an incident of mass mortality following a flash flood.[137]
Dispute[]
Scientists such as Alan Feduccia have cited Protoavis in an attempt to refute the hypothesis that birds evolved from dinosaurs.[116] However, the only consequence would be to push back the point of divergence further back in time.[138] At the time when such claims were originally made, the affiliation of birds and maniraptoran theropods which today is well-supported and generally accepted by most ornithologists was much more contentious; most Mesozoic birds have only been discovered since then. Chatterjee himself has since used Protoavis to support a close relationship between dinosaurs and birds.[139]
"As there remains no compelling data to support the avian status of Protoavis or taxonomic validity thereof, it seems mystifying that the matter should be so contentious. The author very much agrees with Chiappe in arguing that at present, Protoavis is irrelevant to the phylogenetic reconstruction of Aves. While further material from the Dockum beds may vindicate this peculiar archosaur, for the time being, the case for Protoavis is non-existent."[138]
Claimed temporal paradox[]
The temporal paradox, or time problem is a controversial issue in the evolutionary relationships of feathered dinosaurs and birds. It was originally conceived of by paleornithologist Alan Feduccia.[116][140] The concept is based on the apparent following facts. The consensus view is that birds evolved from dinosaurs, but the most bird-like dinosaurs, and those most closely related to birds (the maniraptorans), are known mostly from the Cretaceous, by which time birds had already evolved and diversified. If bird-like dinosaurs are the ancestors of birds they should be older than birds, but Archaeopteryx is 155 million years old, while the very bird-like Deinonychus is 35 million years younger. This idea is sometimes summarized as "you can't be your own grandmother".[141] The development of avian characteristics in dinosaurs supposedly should have led to the first modern bird appearing about 60 million years ago. However, Archaeopteryx lived 150 million years ago, long before any of the bird changes took place in dinosaurs. Each of the feathered dinosaur families developed avian-like features in its own way. Thus there were many several different lines of evolution. Archaeopteryx was merely the result of one such line.
Numerous researchers have discredited the idea of the temporal paradox. Witmer (2002) summarized this critical literature by pointing out that there are at least three lines of evidence that contradict it.[118] First, no one has proposed that maniraptoran dinosaurs of the Cretaceous are the ancestors of birds. They have merely found that dinosaurs like dromaeosaurs, troodontids and oviraptorosaurs are close relatives of birds. The true ancestors are thought to be older than Archaeopteryx, perhaps Early Jurassic or even older. The scarcity of maniraptoran fossils from this time is not surprising, since fossilization is a rare event requiring special circumstances and fossils may never be found of animals in sediments from ages that they actually inhabited. Secondly, fragmentary remains of maniraptoran dinosaurs actually have been known from Jurassic deposits in China, North America, and Europe for many years. The femur of a tiny maniraptoran from the Late Jurassic of Colorado was reported by Padian and Jensen in 1989.[142] In a 2009 article in the journal Acta Palaeontologica Polonica, six velociraptorine dromaeosaurid teeth were described as being recovered from a bone bed in Langenberg Quarry of Oker (Goslar, Germany). These teeth are notable in this instance in that they dated back to the Kimmeridgian stage of the Late Jurassic, roughly 155-150 Ma, and represent some of the earliest dromaeosaurids known to science,[143] further refuting a "temporal paradox". Furthermore, a small, as of yet undescribed troodontid known as WDC DML 001, was announced in 2003 as being found in the Late Jurassic Morrison Formation of eastern/central Wyoming.[144] The presence of this derived maniraptoran in Jurassic sediments is a strong refutation of the "temporal paradox".[144] Third, if the temporal paradox would indicate that birds should not have evolved from dinosaurs, then what animals are more likely ancestors considering their age? Brochu and Norell (2001) analyzed this question using several of the other archosaurs that have been proposed as bird ancestors, and found that all of them create temporal paradoxes—long stretches between the ancestor and Archaeopteryx where there are no intermediate fossils—that are actually worse.[145] Thus, even if one used the logic of the temporal paradox, one should still prefer dinosaurs as the ancestors to birds.
Quick & Ruben (2009)[]


{kind=link}
Alleged diaphragm of Sinosauropteryx highlighted.[146][147]

{kind=link}
Fossil and drawing of Sinosauropteryx prima, showing protofeathers along the dorsal side and tail.
In Quick & Ruben's 2009 paper, they argue that modern birds are fundamentally different from non-avian dinosaurs in terms of abdominal soft-tissue morphology, and therefore birds cannot be modified dinosaurs.[148] The paper asserts that a specialized 'femoral-thigh complex', combined with a synsacrum and ventrally separated pubic bones, provides crucial mechanical support for the abdominal wall in modern birds, and has thereby allowed the evolution of large abdominal air-sacs that function in respiration. In contrast, say the authors, theropod dinosaurs lack these features and had a highly mobile femur that cannot have been incorporated into abdominal support. Therefore, non-avian theropods cannot have had abdominal air-sacs that functioned like those of modern birds, and non-avian theropods were fundamentally different from modern birds. However, this was not mentioned in the paper, but was of course played-up in the press interviews.[149][150] The paper never really demonstrate anything, but merely try to shoot holes in a given line of supporting evidence. It has been argued that respiratory turbinates supposedly falsify dinosaur endothermy,[151] even though it has never been demonstrated that respiratory turbinates really are a requirement for any given physiological regime, and even though there are endotherms that lack respiratory turbinates. The innards of Sinosauropteryx and Scipionyx also supposedly falsify avian-like air-sac systems in non-avian coelurosaurs and demonstrate a crocodilian-like hepatic piston diaphragm,[146][147] even though personal interpretation is required to accept that this claim might be correct. Furthermore, even though crocodilians and dinosaurs are fundamentally different in pelvic anatomy, some living birds have the key soft-tissue traits reported by Ruben et al. in Sinosauropteryx and Scipionyx, and yet still have an avian respiratory system. For a more detailed rebuttal of Quick & Ruben's paper, see this post by Darren Naish at Tetrapod Zoology.
Genuine feathers?[]
There have been claims that the supposed feathers of the Chinese fossils are a preservation artifact.[42] Despite doubts, the fossil feathers have roughly the same appearance as those of birds fossilized in the same locality, so there is no serious reason to think they are of different nature; moreover, no non-theropod fossil from the same site shows such an artifact, but sometimes show unambiguous hair (some mammals) or scales (some reptiles).[92][152][153]
Sinosauropteryx[]
- See also: Feathered dinosaurs#Sinosauropteryx 2
Some researchers have interpreted the filamentous impressions around Sinosauropteryx fossils as remains of collagen fibers, rather than primitive feathers. Since they are clearly external to the body, these researchers have proposed that the fibers formed a frill on the back of the animal and underside of its tail, similar to some modern aquatic lizards.[154]
This would refute the proposal that Sinosauropteryx is the most basal known theropod genus with feathers, and also questions the current theory of feather origins itself. It calls into question the idea that the first feathers evolved not for flight but for insulation, and that they made their first appearance in relatively basal dinosaur lineages that later evolved into modern birds. [155]
The Archaeoraptor fake[]
- Main article: Archaeoraptor
In 1999, a supposed 'missing link' fossil of an apparently feathered dinosaur named "Archaeoraptor liaoningensis", found in Liaoning Province, northeastern China, turned out to be a forgery. Comparing the photograph of the specimen with another find, Chinese paleontologist Xu Xing came to the conclusion that it was composed of two portions of different fossil animals. His claim made National Geographic review their research and they too came to the same conclusion.[156] The bottom portion of the "Archaeoraptor" composite came from a legitimate feathered dromaeosaurid now known as Microraptor,[157] and the upper portion from a previously-known primitive bird called Yanornis.[158]
Flying and gliding[]
[]


{kind=link}
Rahonavis ostromi, a dromaeosaurid originally classified as an avialan bird, may have been capable of flight.

{kind=link}
Artist's rendition of Microraptor.
The ability to fly or glide has been suggested for at least two dromaeosaurid species. The first, Rahonavis ostromi (originally classified as avian bird, but found to be a dromaeosaurid in later studies)[159][160] may have been capable of powered flight, as indicated by its long forelimbs with evidence of quill knob attachments for long sturdy flight feathers.[161] The forelimbs of Rahonavis were more powerfully built than Archaeopteryx, and show evidence that they bore strong ligament attachments necessary for flapping flight. Luis Chiappe concluded that, given these adaptations, Rahonavis could probably fly but would have been more clumsy in the air than modern birds.[162]
Microraptor[]
Another species of dromaeosaurid, Microraptor gui, may have been capable of gliding using its well-developed wings on both the fore and hind limbs. Microraptor was among the first non-avian dinosaurs discovered with the impressions of feathers and wings. On Microraptor, the long feathers on the forelimbs possess asymmetrical vanes. The external vanes are narrow, while the internal ones are broad. In addition, Microraptor possessed elongated remiges with asymmetrical vanes that demonstrate aerodynamic function on the hind limbs.[7][43][163][164] A 2005 study by Sankar Chatterjee suggested that the wings of Microraptor functioned like a split-level "biplane", and that it likely employed a phugoid style of gliding, in which it would launch from a perch and swoop downward in a 'U' shaped curve, then lift again to land on another tree, with the tail and hind wings helping to control its position and speed.[165] Chatterjee also found that Microraptor had the basic requirements to sustain level powered flight in addition to gliding.[166]
Microraptor had two sets of wings, on both its forelegs and hind legs. The long feathers on the legs of Microraptor were true flight feathers as seen in modern birds, with asymmetrical vanes on the arm, leg, and tail feathers.[78][166] As in bird wings, Microraptor had both primary (anchored to the hand) and secondary (anchored to the arm) flight feathers.[78] This standard wing pattern was mirrored on the hind legs, with flight feathers anchored to the upper foot bones as well as the upper and lower leg.[166] It had been proposed by Chinese scientists that the animal glided and probably lived in trees, pointing to the fact that wings anchored to the feet of Microraptor would have hindered their ability to run on the ground, and suggest that all primitive dromaeosaurids may have been arboreal.[7]


{kind=link}
Microraptor specimen with feather impressions.
Sankar Chatterjee determined in 2005 that, in order for the creature to glide or fly, the wings must have been on different levels (as on a biplane) and not overlaid (as on a dragonfly), and that the latter posture would have been anatomically impossible.[165] Using this biplane model, Chatterjee was able to calculate possible methods of gliding, and determined that Microraptor most likely employed a phugoid style of gliding—launching itself from a perch, the animal would have swooped downward in a deep 'U' shaped curve and then lifted again to land on another tree. The feathers not directly employed in the biplane wing structure, like those on the tibia and the tail, could have been used to control drag and alter the flight path, trajectory, etc. The orientation of the hind wings would also have helped the animal control its gliding flight.[165] In 2007, Chatterjee used computer algorithms that test animal flight capacity to determine whether or not Microraptor was capable of true, powered flight, in addition to passive gliding. The resulting data showed that Microraptor did have the requirements to sustain level powered flight, so it is theoretically possible that the animal flew on occasion in addition to gliding.[166]
Saurischian integumentary structures[]
- For a full list of dinosaurs discovered with evidence of integumentary structures, see #List of dinosaur genera preserved with evidence of feathers.
The hip structure possessed by modern birds actually evolved independently from the "lizard-hipped" saurischians (specifically, a sub-group of saurischians called the Maniraptora) in the Jurassic Period. In this example of convergent evolution, birds developed hips oriented similar to the earlier ornithischian hip anatomy, in both cases possibly as an adaptation to a herbivorous or omnivorous diet.
In Saurischia, maniraptorans are characterized by long arms and three-fingered hands, as well as a "half-moon shaped" (semi-lunate) bone in the wrist (carpus). Maniraptorans are the only dinosaurs known to have breast bones (ossified sternal plates). In 2004, Tom Holtz and Halszka Osmólska pointed out six other maniraptoran characters relating to specific details of the skeleton. Unlike most other saurischian dinosaurs, which have pubic bones that point forward, several groups of maniraptorans have an ornithischian-like backwards-pointing hip bone. A backward-pointing hip characterizes the therizinosaurs, dromaeosaurids, avialans, and some primitive troodontids. The fact that the backward-pointing hip is present in so many diverse maniraptoran groups has led most scientists to conclude that the "primitive" forward-pointing hip seen in advanced troodontids and oviraptorosaurs is an evolutionary reversal, and that these groups evolved from ancestors with backward-pointing hips.[167]
Modern pennaceous feathers and remiges are known from advanced maniraptoran groups (Oviraptorosauria and Paraves). More primitive maniraptorans, such as therizinosaurs (specifically Beipiaosaurus), preserve a combination of simple downy filaments and unique elongated quills.[37][168] Powered and/or gliding flight is present in members of Avialae, and possibly in some dromaeosaurids such as Rahonavis and Microraptor.[162] Simple feathers are known from more primitive coelurosaurs such as Sinosauropteryx, and possibly from even more distantly related species such as the ornithischian Tianyulong and the flying pterosaurs. Thus it appears as if some form of feathers or down-like integument would have been present in all maniraptorans, at least when they were young.[169]
Therizinosaurs[]

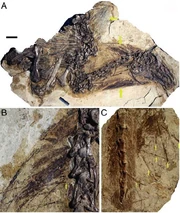
{kind=link}
New fossil specimen of Beipiaosaurus showing preserved filaments.


{kind=link}
A reconstruction of Beipiaosaurus shows how the dinosaur may have displayed its feathers.
Skin impressions from the type specimen of Beipiaosaurus inexpectus indicated that the body was covered predominately by downy feather-like fibers, similar to those of Sinosauropteryx, but longer, and are oriented perpendicular to the arm. Xu et al., who described the specimen, suggested that these downy feathers represent an intermediate stage between Sinosauropteryx and more advanced birds (Avialae).[37]
Unique among known theropods, Beipiaosaurus also possessed a secondary coat of much longer, simpler feathers that rose out of the down layer.[170] These unique feathers (known as EBFFs, or elongated broad filamentous feathers) were first described by Xu et al. in 2009, based on a specimen consisting of the torso, head and neck. Xu and his team also found EBFFs in the original type specimen of B. inexpectus, revealed by further preparation.[168] The holotype also preserved a pygostyle-like structure.[171] The holotype was discovered in two phases. Limb fragments and dorsal and cervical vertebrae were discovered initially. The discovery site was re-excavated later on, and this time an articulated tail and partial pelvis were discovered. All come from the same individual.[171]
The holotype has the largest proto-feathers known of any feathered dinosaur, with the author and paleontologist Xing Xu stating: "Most integumentary filaments are about 50 mm in length, although the longest is up to 70 mm. Some have indications of branching distal ends.".[37] The holotype also had preserved dense patches of parallel integumentary structures in association with its lower arm and leg.[37]
Thick, stiff, spine-like structures were recovered sprouting from the new specimen's throat region, the back of its head, its neck and its back. New preparation of the holotype reveals that the same structures are also present on the tail (though not associated with the pygostyle-like structure).[168]
{kind=link}
Part of the jumbled holotype specimen of Beipiaosaurus on the right. Close-up of tail tip at left. Both show evidence of integumentary structures.
The EBFFs differ from other feather types in that they consist of a single, unbranched filament. Most other primitive feathered dinosaurs have down-like feathers made up of two or more filaments branching out from a common base or along a central shaft. The EBFFs of Beipiaosaurus are also much longer than other primitive feather types, measuring about 100-150 millimeters (4-6 inches) long, roughly half the length of the neck. In Sinosauropteryx, the longest feathers are only about 15% of the neck length. The EBFFs of Beipiaosaurus are also unusually broad, up to 3 mm wide in the type specimen. The broadest feathers of Sinosauropteryx are only 0.2 mm wide, and only slightly wider in larger forms such as Dilong. Additionally, where most primitive feather types are circular in cross section, EBFFs appear to be oval-shaped. None of the preserved EBFFs were curved or bent beyond a broad arc in either specimen, indicating that they were fairly stiff. They were probably hollow, at least at the base.[168]
In a 2009 interview, Xu stated: "Both [feather types] are definitely not for flight, inferring the function of some structures of extinct animals would be very difficult, and in this case, we are not quite sure whether these feathers are for display or some other functions." He speculated that the finer feathers served as an insulatory coat and that the larger feathers were ornamental, perhaps for social interactions such as mating or communication.[170]
Tyrannosauroids[]


{kind=link}
Guanlong may have had a coat of primitive feathers.
Long filamentous structures have been preserved along with skeletal remains of numerous coelurosaurs from the Early Cretaceous Yixian Formation and other nearby geological formations from Liaoning, China.[41] These filaments have usually been interpreted as "protofeathers," homologous with the branched feathers found in birds and some non-avian theropods,[32][172] although other hypotheses have been proposed.[33] A skeleton of Dilong was described in the scientific journal Nature in 2004 that included the first example of "protofeathers" in a tyrannosauroid from the Yixian Formation of China. Similarly to down feathers of modern birds, the "protofeathers" found in Dilong were branched but not pennaceous, and may have been used for insulation.[38]

{kind=link}
Dilong, a basal tyrannosauroid, was covered in protofeathers (shown).

{kind=link}
Dilong, one of the earliest and most primitive known tyrannosauroids, had a covering of simple feathers or protofeathers, and scales.
The presence of "protofeathers" in basal tyrannosauroids is not surprising, since they are now known to be characteristic of coelurosaurs, found in other basal genera like Sinosauropteryx,[32] as well as all more derived groups.[41] Rare fossilized skin impressions of large tyrannosaurids lack feathers, however, instead showing skin covered in scales.[173][174] While it is possible that protofeathers existed on parts of the body which have not been preserved, a lack of insulatory body covering is consistent with modern multi-ton mammals such as elephants, hippopotamuses, and most species of rhinoceros. Alternatively, secondary loss of "protofeathers" in large tyrannosaurids may be analogous with the similar loss of hair in the largest modern mammals like elephants, where a low surface area-to-volume ratio slows down heat transfer, making insulation by a coat of hair unnecessary.[38] Therefore, as large animals evolve in or disperse into warm climates, a coat of fur or feathers loses its selective advantage for thermal insulation and can instead become a disadvantage, as the insulation traps excess heat inside the body, possibly overheating the animal. Protofeathers may also have been secondarily lost during the evolution of large tyrannosaurids, especially in warm Cretaceous climates.[38] Tyrannosaurus at one stage of its life may have been covered in down-like feathers,[175] although there is no direct fossil evidence of this.[176]
Troodontids[]


{kind=link}
The troodontid Jinfengopteryx elegans was originally classified as a bird related to Archaeopteryx.
A few troodont fossils, including specimens of Mei and Sinornithoides, demonstrate that these animals roosted like birds, with their heads tucked under their forelimbs.[67] These fossils, as well as numerous skeletal similarities to birds and related feathered dinosaurs, support the idea that troodontids probably bore a bird-like feathered coat. The discovery of a fully-feathered, primitive troodontid (Jinfengopteryx) lends support to this. The type specimen of Jinfengopteryx elegans is 55 cm long and from the Qiaotou Formation of Liaoning Province, China.
Troodontids are important to research on the origin of birds because they share many anatomical characters with early birds. Crucially, the substantially complete fossil identified as WDC DML 001 ("Lori"), is a troodontid from the Late Jurassic Morrison Formation, close to the time of Archaeopteryx.[144] The discovery of this Jurassic troodont is positive physical evidence that derived deinonychosaurs were present very near the time that birds arose, and basal paravians must have evolved much earlier.[144] This fact strongly invalidates the "temporal paradox" cited by the few remaining opponents of the idea that birds are closely related to dinosaurs. (see claimed temporal paradox below.)
Dromaeosaurids[]


{kind=link}
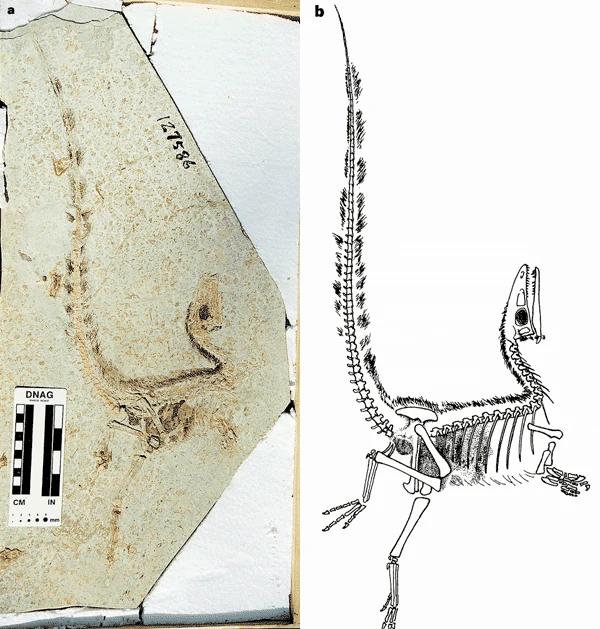
Fossil of Sinornithosaurus millenii, the first evidence of feathers in dromaeosaurids.
There is a large body of evidence showing that dromaeosaurids were covered in feathers. Some dromaeosaurid fossils preserve long, pennaceous feathers on the hands and arms (remiges) and tail (rectrices), as well as shorter, down-like feathers covering the body.[177][172] Other fossils, which do not preserve actual impressions of feathers, still preserve the associated bumps on the forearm bones where long wing feathers would have attached in life.[178] Overall, this feather pattern looks very much like Archaeopteryx.[172]
The first known dromaeosaur with definitive evidence of feathers was Sinornithosaurus, reported from China by Xu et al. in 1999.[177] NGMC 91-A, the Sinornithosaurus-like theropod informally dubbed "Dave", possessed unbranched fibers in additional to more complex branched and tufted structures.[179] Many other dromaeosaurid fossils have been found with feathers covering their bodies, some with fully-developed feathered wings. Several even show evidence of a second pair of wings on the hind legs, including Microraptor and Cryptovolans.[7] While direct feather impressions are only possible in fine-grained sediments, some fossils found in coarser rocks show evidence of feathers by the presence of quill knobs, the attachment points for wing feathers possessed by some birds. The dromaeosaurids Rahonavis and Velociraptor have both been found with quill knobs, showing that these forms had feathers despite no impressions having been found. In light of this, it is most likely that even the larger ground-dwelling dromaeosaurids bore feathers, since even flightless birds today retain most of their plumage, and relatively large dromaeosaurids, like Velociraptor, are known to have retained pennaceous feathers.[45][178] Though some scientists had suggested that the larger dromaeosaurids lost some or all of their insulatory covering, the discovery of feathers in Velociraptor specimens has been cited as evidence that all members of the family retained feathers.[92][178]
Velociraptor[]
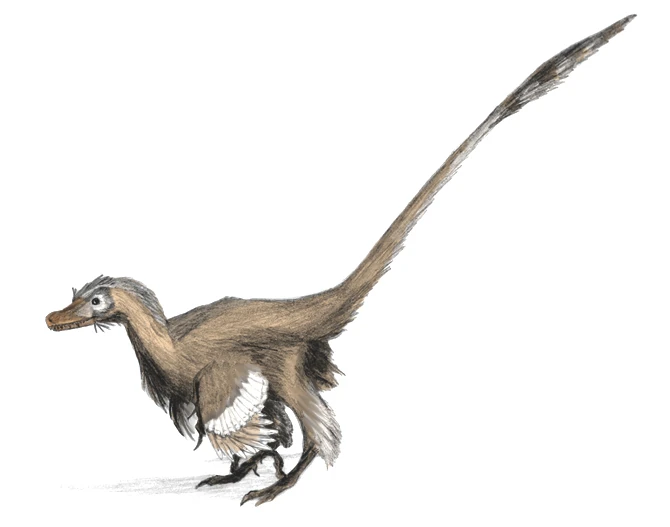

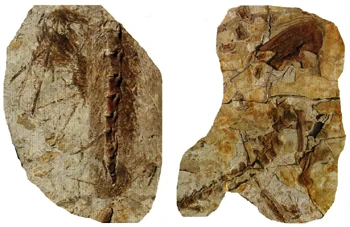
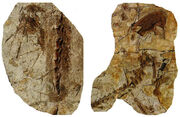
{kind=link}
Velociraptor mongoliensis restored with large wing feathers, as evidenced by fossil quill knobs.
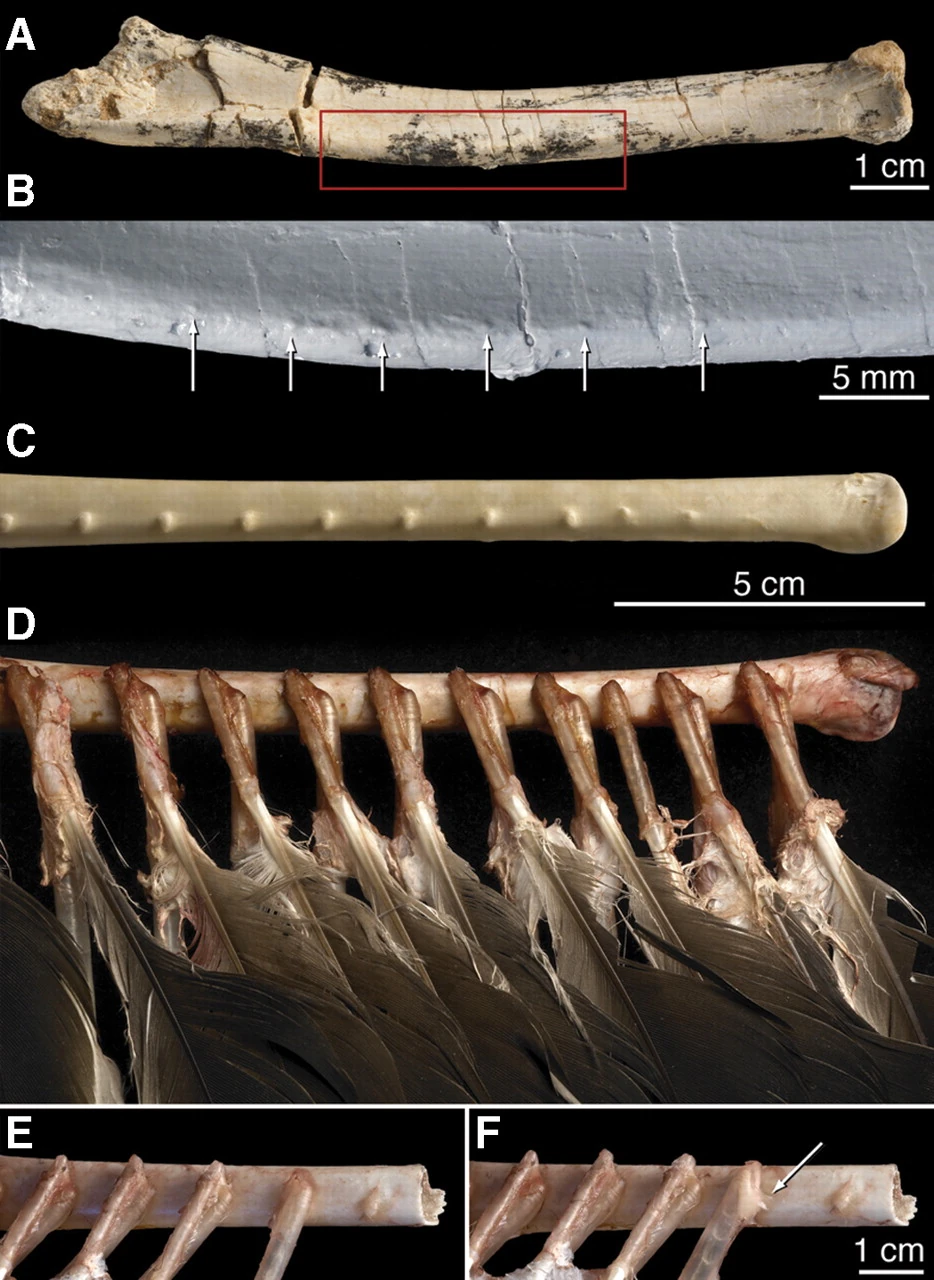
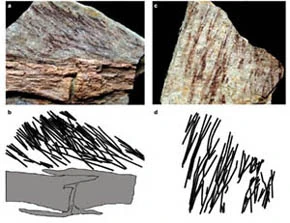
{kind=link}
Right ulna of Velociraptor IGM 100/981 clearly showing six evenly spaced feather quill knobs. Right ulna of a turkey vulture for comparison.[178]
Fossils of dromaeosaurids more primitive than Velociraptor are known to have had feathers covering their bodies, and fully developed, feathered wings.[7] The fact that the ancestors of Velociraptor were feathered and possibly capable of flight long suggested to paleontologists that Velociraptor bore feathers as well, since even flightless birds today retain most of their feathers.
In September 2007, Alan Turner, Peter Makovicky, and Mark Norell reported the presence of quill knobs on the ulna of a Velociraptor specimen from Mongolia.[178] Fourteen bumps approximately 4mm apart were found in a straight line along the bone, directly corresponding to the same structures in living birds, the bumps serving as an anchor for the secondary feathers. These bumps on bird wing bones show where feathers anchor, and their presence on Velociraptor indicate it too had feathers. According to paleontologist Alan Turner,
A lack of quill knobs does not necessarily mean that a dinosaur did not have feathers. Finding quill knobs on Velociraptor, though, means that it definitely had feathers. This is something we'd long suspected, but no one had been able to prove.[180]
Co-author Mark Norell, Curator-in-Charge of fossil reptiles, amphibians and birds at the American Museum of Natural History, also weighed in on the discovery, saying:
The more that we learn about these animals the more we find that there is basically no difference between birds and their closely related dinosaur ancestors like velociraptor. Both have wishbones, brooded their nests, possess hollow bones, and were covered in feathers. If animals like velociraptor were alive today our first impression would be that they were just very unusual looking birds.[180]


{kind=link}
Illustration of a feathered Velociraptor making a trackway.
According to Turner and co-authors Norell and Peter Makovicky, quill knobs are not found in all prehistoric birds, and their absence does not mean that an animal was not feathered – flamingos, for example, have no quill knobs. However, their presence confirms that Velociraptor bore modern-style wing feathers, with a rachis and vane formed by barbs. The forearm specimen on which the quill knobs were found (specimen number IGM 100/981) represents an animal 1.5 meters in length (5 ft) and 15 kilograms (33 lbs) in weight. Based on the spacing of the six preserved knobs in this specimen, the authors suggested that Velociraptor bore 14 secondaries (wing feathers stemming from the forearm), compared with 12 or more in Archaeopteryx, 18 in Microraptor, and 10 in Rahonavis. This type of variation in the number of wing feathers between closely related species, the authors asserted, is to be expected, given similar variation among modern birds.
Turner and colleagues interpreted the presence of feathers on Velociraptor as evidence against the idea that the larger, flightless maniraptorans lost their feathers secondarily due to larger body size. Furthermore, they noted that quill knobs are almost never found in flightless bird species today, and that their presence in Velociraptor (presumed to have been flightless due to its relatively large size and short forelimbs) is evidence that the ancestors of dromaeosaurids could fly, making Velociraptor and other large members of this family secondarily flightless, though it is possible the large wing feathers inferred in the ancestors of Velociraptor had a purpose other than flight. The feathers of the flightless Velociraptor may have been used for display, for covering their nests while brooding, or for added speed and thrust when running up inclined slopes.[178]
Sinornithosaurus[]
The preserved impressions of integumentary structures in Sinornithosaurus were composed of filaments, and showed two features that indicate they are early feathers. First, several filaments were joined together into "tufts", similar to the way down is structured. Second, a row of filaments (barbs) were joined together to a main shaft (rachis), making them similar in structure to normal bird feathers. However, they do not have the secondary branching and tiny little hooks (barbules) that modern feathers have, which allow the feathers of modern birds to form a discrete vane. The filaments are arranged in a parallel fashion to each other, and are perpendicular to the bones.[87] In specimen NGMC - 91, the feathers covered the entire body, including the head in front of the eye, the neck, wing - like sprays on the arms, long feathers on the thighs, and a lozenge - shaped fan on the tail like that of Archaeopteryx.[179]
Pedopenna[]

{kind=link}
Artist's restoration of Pedopenna.
Pedopenna is a maniraptoran theropod that shows evidence of avian affinities that are further evidence of the dinosaur-bird evolutionary relationship. Apart from having a very bird-like skeletal structure in its legs, Pedopenna was remarkable due to the presence of long pennaceous feathers on the metatarsus (foot).[181] Some deinonychosaurs are also known to have these 'hind wings', but those of Pedopenna differ from those of animals like Microraptor.[181] Pedopenna hind wings were smaller and more rounded in shape. The longest feathers were slightly shorter than the metatarsus, at about 55 mm (2 in) long.[181] Additionally, the feathers of Pedopenna were symmetrical, unlike the asymmetrical feathers of some deinonychosaurs and birds. Since asymmetrical feathers are typical of animals adapted to flying, it is likely that Pedopenna represents an early stage in the development of these structures.[181] While many of the feather impressions in the fossil are weak, it is clear that each possessed a rachis and barbs, and while the exact number of foot feathers is uncertain, they are more numerous than in the hind-wings of Microraptor.[181] Pedopenna also shows evidence of shorter feathers overlying the long foot feathers, evidence for the presence of coverts as seen in modern birds. Since the feathers show fewer aerodynamic adaptations than the similar hind wings of Microraptor, and appear to be less stiff, suggests that if they did have some kind of aerodynamic function, it was much weaker than in deinonychosaurs and birds.[181] Xu and Zhang, in their 2005 description of Pedopenna, suggested that the feathers could be ornamental, or even vestigial. It is possible that a hind wing was present in the ancestors of deinonychosaurs and birds, and later lost in the bird lineage, with Pedopenna representing an intermediate stage where the hind wings are being reduced from a functional gliding apparatus to a display or insulatory function.[181]
Avialans and compsognathids[]
Anchiornis[]
Anchiornis is notable for its proportionally long forelimbs, which measured 80% of the total length of the hind limbs. This is similar to the condition in early avians such as Archaeopteryx, and the authors pointed out that long forelimbs are necessary for flight. It is possible that Anchiornis was able to fly or glide, and may have had a functional airfoil. Anchiornis also had a more avian wrist than other non-avian theropods. Anchiornis has hind leg proportions more like those of lower theropod dinosaurs than avialans. Faint, carbonized feather impressions were preserved in patches in the type specimen. Feathers on the torso measured an average of 20 mm in length, but the feathers were too poorly preserved to ascertain details of their structure.[182] A cladistic analysis indicated that Anchiornis is part of the avian lineage, but outside of the clade that includes Archaeopteryx and modern birds, strongly suggesting that Anchiornis was a basal member of the Avialae and the sister taxon of Aves.[182] Anchiornis can therefore be considered to be a non-avian avialian.
Sinosauropteryx[]
All specimens of Sinosauropteryx preserve integumentary structures (filaments arising from the skin) which most paleontologists interpret as very primitive feathers. These short, down-like filaments are preserved all along the back half of the skull, arms, neck, back, and top and bottom of the tail. Additional patches of feathers have been identified on the sides of the body, and paleontologist Chen, Dong and Zheng proposed that the density of the feathers on the back and the randomness of the patches elsewhere on the body indicated the animals would have been fully feathered in life, with the ventral feathers having been removed by decomposition.[32]


{kind=link}
An undescribed fossil specimen labeled Sinosauropteryx on display in the Hong Kong Science Museum.


{kind=link}
Holotype fossil of Sinosauropteryx prima with impressions of integumentary filaments.
The filaments are preserved with a gap between the bones, which several authors have noted corresponds closely to the expected amount of skin and muscle tissue that would have been present in life. The feathers adhere close to the bone on the skull and end of the tail, where little to no muscle was present, and the gap increases over the back vertebrae, where more musculature would be expected, indicating that the filaments were external to the skin and do not correspond with sub-cutaneous structures.[32]
The random positioning of the filaments and often "wavy" lines of preservation indicate that they were soft and pliable in life. Examination with microscopes shows that each individual filament appears dark along the edges and light internally, suggesting that they were hollow, like modern feathers. Compared to modern mammals the filaments were quite coarse, with each individual strand much larger and thicker than the corresponding hairs of similarly sized mammals.[32]
The length of the filaments varied across the body. They were shortest just in front of the eyes, with a length of 13 mm. Going further down the body, the filaments rapidly increase in length until reaching 35 mm long over the shoulder blades. The length remains uniform over the back, until beyong the hips, when the filaments lengthen again and reach their maximum length midway down the tail at 40 mm. The filaments on the underside of the tail are shorter overall and decrease in length more rapidly than those on the dorsal surface. By the 25th tail vertebrae, the filaments on the underside reach a length of only 35 mm. The longest feathers present on the forearm measured 14 mm.[32]
Overall, the filaments most closely resemble the "plumules" or down-like feathers of some modern birds, with a very short quill and long, thin barbs. The same structures are seen in other fossils from the Yixian Formation, including Confuciusornis.[32]
Analysis of the fossils of Sinosauropteryx have shown an alternation of lighter and darker bands preserved on the tail, giving us an idea of what the animal looked like in real life. This banding is probably due to preserved areas of melanin, which can produce dark tones in fossils.[183]
Scansoriopterygids[]


{kind=link}
Scansoriopteryx was the first non-avian dinosaur found that had clear adaptations to an arboreal or semi-arboreal lifestyle.[8]
Epidendrosaurus[]
The type specimen of Epidendrosaurus also preserved faint feather impressions at the end of the tail, similar to the pattern found in the dromaeosaurid Microraptor.[8] While the reproductive strategies of Epidendrosaurus itself remain unknown, several tiny fossil eggs discovered in Phu Phok, Thailand (one of which contained the embryo of a theropod dinosaur) may have been laid by a small dinosaur similar to Epidendrosaurus or Microraptor. The authors who described these eggs estimated the dinosaur they belonged to would have had the adult size of a modern Goldfinch.[184]
Epidexipteryx[]
Scansoriopteryx[]
Scansoriopteryx fossils preserve impressions of wispy, down-like feathers around select parts of the body, forming V-shaped patterns similar to those seen in modern down feathers. The most prominent feather impressions trail from the left forearm and hand. The longer feathers in this region led Czerkas and Yuan to speculate that adult scansoriopterygids may have had reasonably well-developed wing feathers which could have aided in leaping or rudimentary gliding, though they ruled out the possibility that Scansoriopteryx could have achieved powered flight. Like other maniraptorans, Scansoriopteryx had a semilunate (half-moon shaped) bone in the wrist that allowed for bird-like folding motion in the hand. Even if powered flight was not possible, this motion could have aided maneuverability in leaping from branch to branch.[185] Scales were also preserved near the base of the tail.[185] For more on the implications of this discovery, see Scansoriopteryx#Implications.
Oviraptorosaurs[]
Oviraptorosaurs, like dromaeosaurs, are so bird-like that several scientists consider them to be true birds, more advanced than Archaeopteryx. Gregory S. Paul has written extensively on this possibility, and Teresa Maryańska and colleagues published a technical paper detailing this idea in 2002.[186][45][15] Michael Benton, in his widely-respected text Vertebrate Palaeontology, also included oviraptorosaurs as an order within the class Aves.[187] However, a number of researchers have disagreed with this classification, retaining oviraptorosaurs as non-avialan maniraptorans slightly more primitive than the dromaeosaurs.[188]
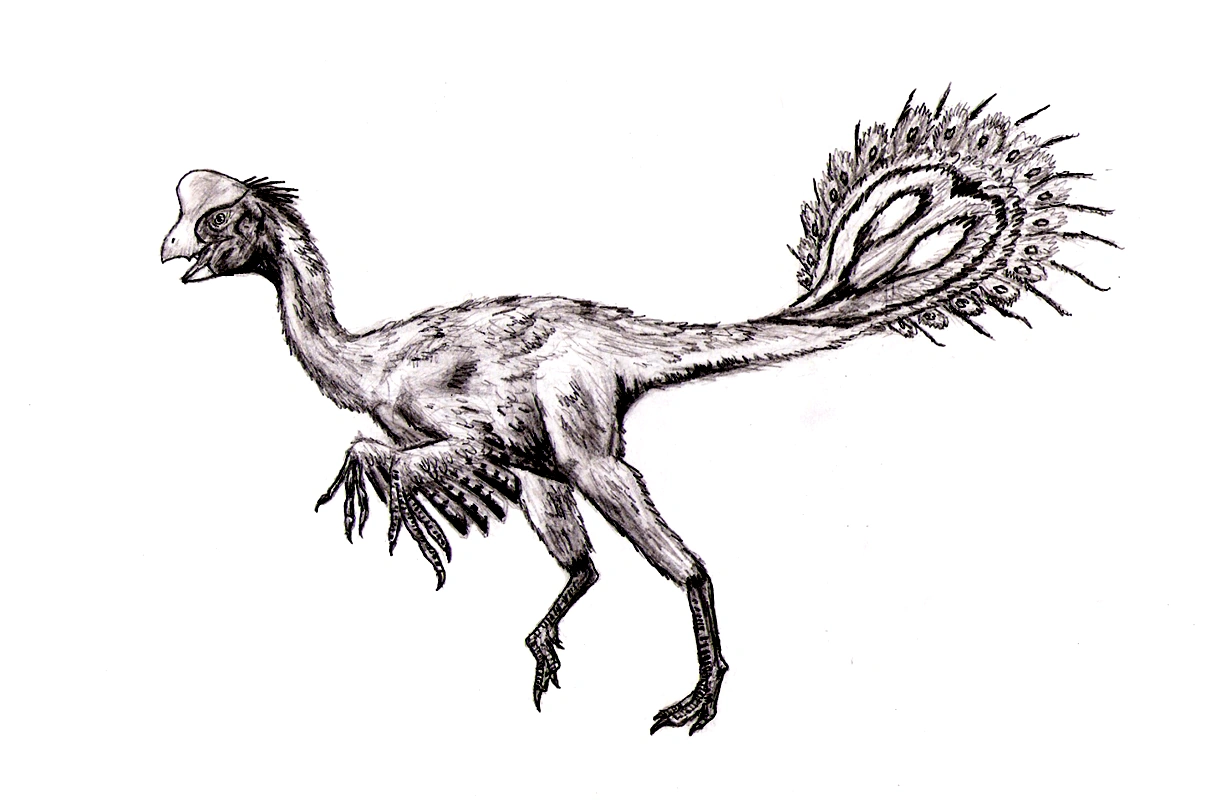
{kind=link}
Nomingia gobiensis, a caenagnathid oviraptorosaur, sported a pygostyle-like mass of fused vertebrae at the tail end that probably supported a feather fan as in Caudipteryx.[189]
Evidence for feathered oviraptorosaurs exists in several forms. Most directly, two species of primitive oviraptorosaurs (Caudipteryx) have been found with impressions of well developed feathers, most notably on the wings and tail, suggesting that they functioned at least partially for display.[190] Secondly, at least one oviraptorosaur (Nomingia) was preserved with a tail ending in something like a pygostyle, a bony structure at the end of the tail that, in modern birds, is used to support a fan of feathers.[189] Similarly, quill knobs (anchor points for wing feathers on the ulna) have been reported in the oviraptorosaurian species, Avimimus portentosus.[191] Additionally, a number of oviraptorid specimens have famously been discovered in a nesting position similar to that of modern birds. The arms of these specimens are positioned in such a way that they could perfectly cover their eggs if they had small wings and a substantial covering of feathers.[192] Protarchaeopteryx, an oviraptorosaur, is well known for its fan-like array of 12 rectricial feathers, but it also seems to have sported simple filament-like structures elsewhere on the tail. Soft and downy feathers are preserved in the chest region and tail base, and are also preserved adjacent to the femora.[34]
The bodies and limbs of oviraptorosaurs are arranged in a bird-like manner, suggesting the presence of feathers on the arms which may have been used for insulating eggs or brooding young.[192] Members of Oviraptoridae possess a quadrate bone that shows particularly avian characteristics, including a pneumatizatized, double-headed structure, the presence of the pterygoid process, and articular fossa for the quadrratojugal.[163][193]
Oviraptorids were probably feathered, since some close relatives were found with feathers preserved (Caudipteryx and possibly Protarchaeopteryx).[34][77] Another finding pointing to this is the discovery in Nomingia of a pygostyle, a bone that results from the fusion of the last tail vertebrae and is responsible in birds to hold a fan of feathers in the tail.[189] Finally, the arm position of the brooding Citipati would have been far more effective if feathers were present to cover the eggs.[45]
Caudipteryx[]


{kind=link}
Caudipteryx arm showing the remiges.
Caudipteryx has clear and unambiguously pennaceous feathers, like modern birds, and several cladistic analyses have consistently recovered it as a nonavian, oviraptorid, dinosaur, it provided, at the time of its description, the clearest and most succinct evidence that birds evolved from dinosaurs. Lawrence Witmer stated:
- "The presence of unambiguous feathers in an unambiguously nonavian theropod has the rhetorical impact of an atomic bomb, rendering any doubt about the theropod relationships of birds ludicrous.”"
However, not all scientists agreed that Caudipteryx was unambiguously non-avian, and some of them continued to doubt that general consensus. Paleornithologist Alan Feduccia sees Caudipteryx as a flightless bird evolving from earlier archosaurian dinosaurs rather than from late theropods.[116] Jones et al. (2000)[194] found that Caudipteryx was a bird based on a mathematical comparison of the body proportions of flightless birds and non-avian theropods. Dyke and Norell (2005) criticized this result for flaws in their mathematical methods, and produced results of their own which supported the opposite conclusion.[117] Other researchers not normally involved in the debate over bird origins, such as Zhou, acknowledged that the true affinities of Caudipteryx were debatable.[195]
Ornithomimosaurs[]
In 1997, filament-like integumentary structures were reported to be present in the Spanish ornithomimosaur Pelecanimimus polyodon. Furthermore, one published life restoration depicts Pelecanimimus as having been covered in the same sort of quill-like structures as are present on Sinosauropteryx and Dilong.[196] However, a brief 1997 report that described soft-tissue mineralization in the Pelecanimimus holotype has been taken by most workers as the definitive last word 'demonstrating' that integumentary fibers were absent from this taxon.[197]
However, the report describing soft-tissue mineralization described soft-tissue preservation seen in one small patch of tissue, and the absence of integument here does not provide much information about the distribution of integument on the live animal. This might explain why a few theropod workers (notably Paul Sereno and Kevin Padian) have continued to indicate the presence of filamentous integumentary structures in Pelecanimimus. Feduccia et al. (2005) argued that Pelecanimimus possessed scaly arms and figured some unusual rhomboidal structures in an effort to demonstrate this.[42] The objects that they illustrate do not resemble scales and it remains to be seen whether they are anything to do with the integument of this dinosaur. A full description/monograph on this dinosaur has yet to be published, which might have more information on this subject.
Ornithischian integumentary structures[]
Psittacosaurus[]

{kind=link}
Psittacosaurus sibiricus exhibiting its tail bristles.


{kind=link}
The holotype of Tianyulong (Zheng et al. 2009:fig. 1a)
The integument, or body covering, of Psittacosaurus is known from a Chinese specimen, which most likely comes from the Yixian Formation of Liaoning. The specimen, which is not yet assigned to any particular species, was illegally exported from China, in violation of Chinese law, but was purchased by a German museum and arrangements are being made to return the specimen to China.[95]
Most of the body was covered in scales. Larger scales were arranged in irregular patterns, with numerous smaller scales occupying the spaces between them, similarly to skin impressions known from other ceratopsians, such as Chasmosaurus.[198] However, a series of what appear to be hollow, tubular bristles, approximately 16 centimeters (6.4 in) long, were also preserved, arranged in a row down the dorsal (upper) surface of the tail.[95][198] However, according to Mayr et al., "[a]t present, there is no convincing evidence which shows these structures to be homologous to the structurally different [feathers and protofeathers] of theropod dinosaurs."[95] As the structures are only found in a single row on the tail, it is unlikely that they were used for thermoregulation, but they may have been useful for communication through some sort of display.[95]
Tianyulong[]
Tianyulong is notable for the row of long, filamentous integumentary structures apparent on the back, tail and neck of the fossil.[199] The similarity of these structures with those found on some derived theropods suggests their homology with feathers and raises the possibility that the earliest dinosaurs and their ancestors were covered with analogous dermal filamentous structures that can be considered as primitive feathers (proto-feathers).[199][200]


{kind=link}
Comparison of the Psittacosaurus "quills" (top), Sinosauropteryx protofeathers (left) and Tianyulong "filaments" (right).
The filamentous integumentary structures are preserved on three areas of the fossil: in one patch just below the neck, another one on the back, and the largest one above the tail.[199] The hollow filaments are parallel to each other and are singular with no evidence of branching.[199] They also appear to be relatively rigid, making them more analogous to the integumentary structures found on the tail of Psittacosaurus[95] than to the proto-feather structures found in avian and non-avian theropods. Among the theropods, the structures in Tianyulong are most similar to the singular unbranched proto-feathers of Sinosauropteryx[109] and Beipiaosaurus.[168] The estimated length of the integumentary structures on the tail is about 60 mm which is seven times the height of a caudal vertebra. Their length and hollow nature argue against of them being subdermal structures such as collagen fibers.[199]
Phylogenetics and homology[]
Such dermal structures have previously been reported only in derived theropods and ornithischians, and their discovery in Tianyulong extends the existence of such structures further down in the phylogenetic tree.[199][201] However, the homology between the ornithischian filaments and the theropods proto-feathers is not obvious. If the homology is supported, the consequence is that the common ancestor of both saurischians and ornithischians were covered by feather-like structures and that groups for which skin impression are known such as the sauropods were only secondarily featherless.[202] If the homology is not supported, it would indicate that these filamentous dermal structures evolved independently in saurischians and ornithischians, as well as in other archosaurs such as the pterosaurs.[202] The authors (in supplementary information to their primary article) noted that discovery of similar filamentous structures in the theropod Beipiaosaurus bolstered the idea that the structures on Tianyulong are homologous with feathers.[199] Both the filaments of Tianyulong and the filaments of Beipiaosaurus were laong, singular, and unbranched. In Beipiaosaurus, however, the filaments were flattened. In Tianyulong, the filaments were round in cross section, and therefore closer in structure to the earliest forms of feathers predicted by developmental models.[199]
Some scientists have argued that other dinosaur proto-feathers are actually fibers of collagen that have come loose from the animals' skins. However, collagen fibers are solid structures; based on the long, hollow nature of the filaments on Tianyulong the authors rejected this explanation.[199]
Current knowledge[]


{kind=link}
Cryptovolans, the first dinosaur known to have asymmetric, primary flight feathers on its legs as well as on its arms.[44]
Sinosauropteryx fossil shwoing the presence of filametous structures.

Fossil cast of NGMC 91, a probable specimen of Sinornithosaurus.
After a century of hypotheses without conclusive evidence, especially well-preserved (and legitimate) fossils of feathered dinosaurs were discovered during the 1990s, and more continue to be found. The fossils were preserved in a lagerstätte — a sedimentary deposit exhibiting remarkable richness and completeness in its fossils — in Liaoning, China. The area had repeatedly been smothered in volcanic ash produced by eruptions in Inner Mongolia 124 million years ago, during the Early Cretaceous Period.[21][22] The fine-grained ash preserved the living organisms that it buried in fine detail. The area was teeming with life, with millions of leaves, angiosperms (the oldest known), insects, fish, frogs, salamanders, mammals, turtles, lizards and crocodilians discovered to date.
The most important discoveries at Liaoning have been a host of feathered dinosaur fossils, with a steady stream of new finds filling in the picture of the dinosaur-bird connection and adding more to theories of the evolutionary development of feathers and flight. Norell et al (2007) reported quill knobs from an ulna of Velociraptor mongoliensis, and these are strongly correlated with large and well-developed secondary feathers.[178]
List of dinosaur genera preserved with evidence of feathers[]
A number of non-avian dinosaurs are now known to have been feathered. Direct evidence of feathers exists for the following genera, listed in the order currently accepted evidence was first published. In all examples, the evidence described consists of feather impressions, except those marked with an asterisk (*), which denotes genera known to have had feathers based on skeletal or chemical evidence, such as the presence of quill knobs.
- Avimimus* (1987)[191][114]:536
- Sinosauropteryx (1996)[31]
- Protarchaeopteryx (1997)[77]
- Caudipteryx (1998)[34]
- Rahonavis* (1998)[161]
- Shuvuuia (1999)[96]
- Sinornithosaurus (1999)[203]
- Beipiaosaurus (1999)[204]
- Microraptor (2000)[78]
- Nomingia* (2000)[189]
- Cryptovolans (2002)[43]
- Scansoriopteryx (2002)[185]
- Epidendrosaurus (2002)[8]
- Psittacosaurus? (2002)[95]
- Yixianosaurus (2003)[205]
- Dilong (2004)[38]
- Pedopenna (2005)[181]
- Jinfengopteryx (2005)[206][188]
- Sinocalliopteryx (2007)[207]
- Velociraptor* (2007)[178]
- Epidexipteryx (2008)[101]
- Anchiornis (2009)[182]
- Tianyulong? (2009)[199]
- Note, filamentous structures in some ornithischian dinosaurs (Psittacosaurus, Tianyulong) and pterosaurs may or may not be homologous with the feathers and protofeathers of theropods.[199][208]
Phylogeny and the inference of feathers in other dinosaurs[]
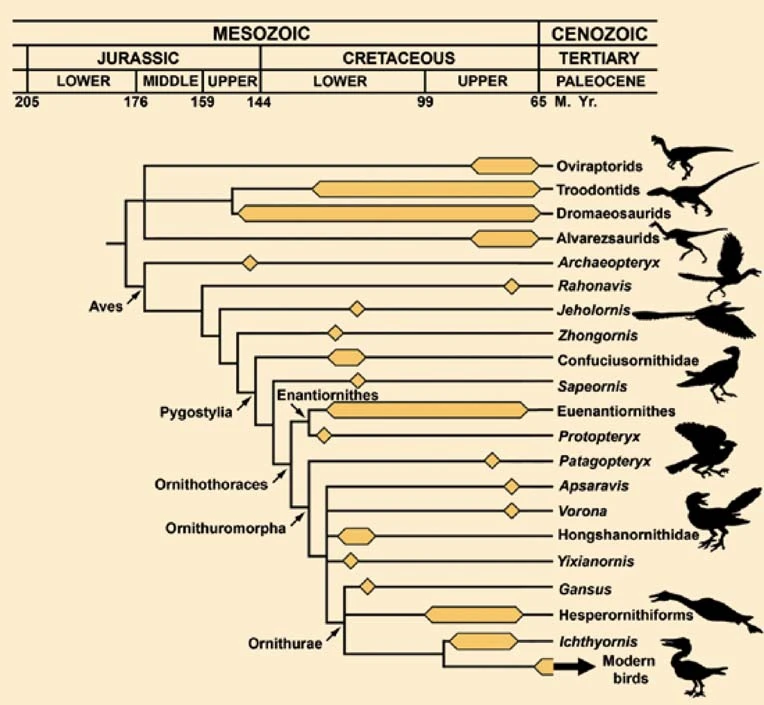
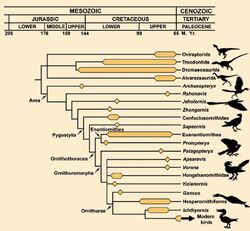
{kind=link}
Cladogram depicting the genealogical relationships among the main lineages of premodern birds and some lineages of nonavian maniraptoran dinosaurs. The known fossil record of these groups is highlighted.


{kind=link}
Anchiornis fills a gap in the transition between the body plans of flying avian birds and non-avian dinosaurs.[182]


{kind=link}
Jinfengopteryx elegans fossil displaying clear evidence of feathers.
Feathered dinosaur fossil finds to date, together with cladistic analysis, suggest that many types of theropod may have had feathers, not just those that are especially similar to birds. In particular, the smaller theropod species may all have had feathers and possibly even the larger theropods (for instance T. rex) may have had feathers, in their early stages of development after hatching.[175] Whereas these smaller animals may have benefited from the insulation of feathers, large adult theropods are unlikely to have had feathers, since inertial heat retention would likely be sufficient to manage heat. Excess internal heat may even have become a problem, had these very large creatures been feathered.
Fossil feather impressions are extremely rare; therefore only a few feathered dinosaurs have been identified so far. However, through a process called phylogenetic bracketing, scientists can infer the presence of feathers on poorly-preserved specimens. All fossil feather specimens have been found to show certain similarities. Due to these similarities and through developmental research almost all scientists agree that feathers could only have evolved once in dinosaurs. Feathers would then have been passed down to all later, more derived species (although it is possible that some lineages lost feathers secondarily). If a dinosaur falls at a point on an evolutionary tree within the known feather-bearing lineages, scientists assume it too had feathers, unless conflicting evidence is found. This technique can also be used to infer the type of feathers a species may have had, since the developmental history of feathers is now reasonably well-known.[92]
Nearly all paleontologists regard birds as coelurosaurian theropod dinosaurs.[124] Within Coelurosauria, multiple cladistic analyses have found support for a clade named Maniraptora, consisting of therizinosauroids, oviraptorosaurs, troodontids, dromaeosaurids, and birds.[209][169][167] Of these, dromaeosaurids and troodontids are usually united in the clade Deinonychosauria, which is a sister group to birds (together forming the node-clade Eumaniraptora) within the stem-clade Paraves.[209][178]
Other studies have proposed alternative phylogenies in which certain groups of dinosaurs that are usually considered non-avian are suggested to have evolved from avian ancestors. For example, a 2002 analysis found oviraptorosaurs to be basal avians.[210] Alvarezsaurids, known from Asia and the Americas, have been variously classified as basal maniraptorans,[209][169][211][212] paravians,[167] the sister taxon of ornithomimosaurs,[213] as well as specialized early birds.[214][215] The genus Rahonavis, originally described as an early bird,[216] has been identified as a non-avian dromaeosaurid in several studies.[178][159] Dromaeosaurids and troodontids themselves have also been suggested to lie within Aves rather than just outside it.[45]:472[217]
The scientists who described the (apparently unfeathered) Juravenator performed a genealogical study of coelurosaurs, including distribution of various feather types. Based on the placement of feathered species in relation to those that have not been found with any type of skin impressions, they were able to infer the presence of feathers in certain dinosaur groups. The following simplified cladogram follows these results, and shows the likely distribution of plumaceous (downy) and pennaceous (vaned) feathers among theropods.[218] Note that the authors inferred pennaceous feathers for Velociraptor based on phylogenetic bracketing, a prediction later confirmed by fossil evidence.[178]
Cladogram[]
| Coelurosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
See also[]
- Origin of birds
- Evolution of birds
- Origin of avian flight
- Birds Came First
- Alan Feduccia
- George Olshevsky
Notes[]
- ^ All known dromaeosaurs have pennaceous feathers on the arms and tail, and substantially thick coat of feathers on the body, especially the neck and breast. Clear fossil evidence of modern avian-style feathers exists for several related dromaeosaurids, including Velociraptor and Microraptor, though no direct evidence is yet known for Deinonychus itself.
- ^ On page 155 of Dinosaurs of the Air by Gregory Paul, there are an accumulated total of 305 potential synapomorphies with birds for all non-avian theropod nodes, 347 for all non-avian dinosauromorph nodes.
Shared features between birds and dinosaurs include:
- A pubis (one of the three bones making up the vertebrate pelvis) shifted from an anterior to a more posterior orientation (see Saurischia), and bearing a small distal "boot".
- Elongated arms and forelimbs and clawed manus (hands).
- Large orbits (eye openings in the skull).
- Flexible wrist with a semi-lunate carpal (wrist bone).
- Double-condyled dorsal joint on the quadrate bone.
- Ossified ucinate process of the ribs.
- Most of the sternum is ossified.
- Broad sternal plates.
- Ossified sternal ribs.
- Brain enlarged above reptilian maximum.
- Overlapping field of vision.
- Olfaction sense reduced.
- An arm/leg length ratio between 0.5 and 1.0
- Lateral exposition of the glenoid in the humeral joint.
- Hollow, thin-walled bones.
- 3-fingered opposable grasping manus (hand), 4-toed pes (foot); but supported by 3 main toes.
- Fused carpometacarpus.
- Metacarpal III bowed posterolaterally.
- Flexibilty of digit III reduced.
- Digit III tightly appressed to digit II.
- Well developed arm folding mechanism.
- Reduced, posteriorly stiffened tail.
- Distal tail stiffened.
- Tail base hyperflexible, especially dorsally.
- Elongated metatarsals (bones of the feet between the ankle and toes).
- S-shaped curved neck.
- Erect, digitgrade (ankle held well off the ground) stance with feet postitioned directly below the body.
- Similar eggshell microstructure.
- Teeth with a constriction between the root and the crown.
- Functional basis for wing power stroke present in arms and pectoral girdle (during motion, the arms were swung down and forward, then up and backwards, describing a "figure-eight" when viewed laterally).
- Expanded pneumatic sinuses in the skull.
- Five or more vertebrae incorporated into the sacrum (hip).
- Posterior caudal vertebrate fused to form the pygostyle.
- Large, strongly built, and straplike scapula (shoulder blade).
- Scapula blades are horizontal.
- Scapula tip is pointed.
- Acromion process is developed, similar to that in Archaeopteryx.
- Retroverted and long coracoids.
- Strongly flexed and subvertical coracoids relative to the scapula.
- Clavicles (collarbone) fused to form a furcula (wishbone).
- U-shaped furcula.
- Hingelike ankle joint, with movement mostly restricted to the fore-aft plane.
- Secondary bony palate (nostrils open posteriorly in throat).
- Pennaceous feathers in some taxa. Proto-feathers, filaments, and integumenatary structures in others.
- Well-developed, symmetrical arm contour feathers.
- Source 1: Are Birds Really Dinosaurs? Dinobuzz, Current Topics Concerning Dinosaurs. Created 9/27/05. Accessed 7/20/09. Copyright 1994-2009 by the Regents of the University of California, all rights reserved.
- Source 2: Kurochkin, E., N. 2006. Parallel Evolution of Theropod Dinoaurs and Birds. Entomological Review 86 (1), pp. S45-S58. doi:10.1134/S0013873806100046
- Source 3: Paul, Gregory S. (2002). "11". Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. pp. 225-227: Table 11.1. ISBN 978-0801867637.
- ^ Xu Xing suggested that the integumentary features present in some pterosaurs and the ornithischian dinosaur Psittacosaurus may be evidence of this first stage.
- ^ Examples in the fossil record may include Sinosauropteryx, Beipiaosaurus, Dilong, and Sinornithosaurus.
- ^ According to Xu Xing, the stage III is supported by the fact that feather follicles developed after barb ridges, along with the follicle having a unique role in the formation of the rachis.
- ^ See Caudipteryx, Protarchaeopteryx, and Sinornithosaurus.
Xu Xing also noted that while the pennaceous feathers of Microraptor differ from those of Caudipteryx and Protarchaeopteryx due to the aerodynamic functions of its feathers, they still belong together in the same stage because they both "evolved form-stiffening barbules" on their feathers. - ^ Remiges are the large feathers of the forelimbs (singular remex). The large feathers that grow from the tail are termed rectrices (singular rectrix).
References[]
- ^ Darwin, Charles R. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. London: John Murray. p. 502pp. http://darwin-online.org.uk/content/frameset?itemID=F373&viewtype=side&pageseq=16.
- ^ Huxley, Thomas H. (1870). "Further evidence of the affinity between the dinosaurian reptiles and birds". Quarterly Journal of the Geological Society of London 26: 12–31.
- ^ Huxley, Thomas H. (1868). "On the animals which are most nearly intermediate between birds and reptiles". Annals of the Magazine of Natural History 4 (2): 66–75.
- ^ Foster, Michael; Lankester, E. Ray 1898–1903. The scientific memoirs of Thomas Henry Huxley. 4 vols and supplement. London: Macmillan.
- ^ Owen, R. (1863): On the Archaeopteryx of von Meyer, with a description of the fossil remains of a long-tailed species, from the Lithographic Slate of Solenhofen. - Philosophical Transactions of the Royal Society of London, 1863: 33-47. London.
- ^ a b Padian K. and Chiappe LM (1998). The origin and early evolution of birds. Biological Reviews 73: 1-42.
- ^ a b c d e f g h i Xu Xing; Zhou Zhonghe; Wang Xiaolin; Kuang Xuewen; Zhang Fucheng; & Du Xiangke (2003). "Four-winged dinosaurs from China". Nature 421 (6921): 335–340. doi:.
- ^ a b c d Zhang, F., Zhou, Z., Xu, X. & Wang, X. (2002). "A juvenile coelurosaurian theropod from China indicates arboreal habits." Naturwissenschaften, 89(9): 394-398. doi:10.1007 /s00114-002-0353-8.
- ^ Fox, W. (1866). Another new Wealden reptile. Athenaeum 2014, 740.
- ^ Naish, D. (2002). The historical taxonomy of the Lower Cretaceous theropods (Dinosauria) Calamospondylus and Aristosuchus from the Isle of Wight. Proceedings of the Geologists' Association 113, 153-163.
- ^ Swinton, W. E. (1936a). Notes on the osteology of Hypsilophodon, and on the family Hypsilophodontidae. Proceedings of the Zoological Society of London 1936, 555-578.
- ^ Swinton, W. E. (1936b). The dinosaurs of the Isle of Wight. Proceedings of the Geologists' Association 47, 204-220.
- ^ Galton, P. M. (1971a). Hypsilophodon, the cursorial non-arboreal dinosaur. Nature 231, 159-161.
- ^ Galton, P. M. (1971b). The mode of life of Hypsilophodon, the supposedly arboreal ornithopod dinosaur. Lethaia 4, 453-465.
- ^ a b Paul, G.S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster.
- ^ a b Olshevsky, G. (2001a). The birds came first: a scenario for avian origins and early evolution, 1. Dino Press 4, 109-117.
- ^ a b Olshevsky, G. (2001b). The birds came first: a scenario for avian origins and early evolution. Dino Press 5, 106-112.
- ^ a b Ostrom, John H. (1969). "Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana". Bulletin of the Peabody Museum of Natural History 30: 1–165.
- ^ Paul, Gregory S. (2000). "A Quick History of Dinosaur Art". in Paul, Gregory S. (ed.). The Scientific American Book of Dinosaurs. New York: St. Martin's Press. pp. 107–112. ISBN 0-312-26226-4.
- ^ El Pais: El 'escándalo archaeoraptor' José Luis Sanz y Francisco Ortega 16/02/2000 Online, Spanish
- ^ a b Swisher Iii, C.C.; Wang, Y.Q.; Wang, X.L.; Xu, X.; Wang, Y. (2001), "Cretaceous age for the feathered dinosaurs of Liaoning, China", Rise of the Dragon: Readings from Nature on the Chinese Fossil Record: 167, http://books.google.com/books?hl=en, retrieved on 2009-09-02
- ^ a b Swisher, C.C.; Xiaolin, W.; Zhonghe, Z.; Yuanqing, W.; Fan, J.I.N.; Jiangyong, Z.; Xing, X.U.; Fucheng, Z.; et al. (2002), "Further support for a Cretaceous age for the feathered-dinosaur beds of Liaoning, China: %u2026", Chinese Science Bulletin 47 (2): 136–139, http://www.springerlink.com/index/W7724740N2320M80.pdf, retrieved on 2009-09-02
- ^ Sereno, Paul C.; & Rao Chenggang (1992). "Early evolution of avian flight and perching: new evidence from the Lower Cretaceous of China". Science 255 (5046): 845–848. doi:. PMID 17756432.
- ^ Hou Lian-Hai; Zhou Zhonghe; Martin, Larry D.; & Feduccia, Alan (1995). "A beaked bird from the Jurassic of China". Nature 377 (6550): 616–618. doi:.
- ^ Novas, F. E., Puerta, P. F. (1997). New evidence concerning avian origins from the Late Cretaceous of Patagonia. Nature 387:390-392.
- ^ Norell, M. A., Clark, J. M., Makovivky, P. J. (2001). Phylogenetic relationships among coelurosaurian dinosaurs. In: Gauthier, J. A., Gall, L. F., eds. New Perspectives on the Origin and Early Evolution of Birds. Yale University Press, New Haven, pp. 49-67.
- ^ Gatesy, S. M., Dial, K. P. (1996). Locomotor modules and the evolution of avian flight. Evolution 50:331-340.
- ^ Gatesy, S. M. (2001). The evolutionary history of the theropod caudal locomotor module. In: Gauthier, J. A., Gall, L. F., eds. New Perspectives on the Origin and Early Evolution of Birds. Yale University Press, New Haven, pp. 333-350.
- ^ Xu, X. (2002). Deinonychosaurian fossils from the Jehol Group of western Liaoning and the coelurosaurian evolution (Dissertation). Chinese Academy of Sciences, Beijing.
- ^ a b c d e f g h i j k l m n o p q Xu Xing (2006). Feathered dinosaurs from China and the evolution of major avian characters. Integrative Zoology 1:4-11. doi:10.1111/j.1749-4877.2006.00004.x
- ^ a b Ji Qiang; & Ji Shu-an (1996). "On the discovery of the earliest bird fossil in China and the origin of birds". Chinese Geology 233: 30–33.
- ^ a b c d e f g h i Chen Pei-ji; Dong Zhiming; & Zhen Shuo-nan. (1998). "An exceptionally preserved theropod dinosaur from the Yixian Formation of China". Nature 391 (6663): 147–152. doi:.
- ^ a b Lingham-Soliar, Theagarten; Feduccia, Alan; & Wang Xiaolin. (2007). "A new Chinese specimen indicates that ‘protofeathers’ in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres". Proceedings of the Royal Society B: Biological Sciences 274 (1620): 1823–1829. doi:.
- ^ a b c d e f Ji Qiang; Currie, Philip J.; Norell, Mark A.; & Ji Shu-an. (1998). "Two feathered dinosaurs from northeastern China". Nature 393 (6687): 753–761. doi:.
- ^ Sloan, Christopher P. (1999). "Feathers for T. rex?". National Geographic 196 (5): 98–107.
- ^ Monastersky, Richard (2000). "All mixed up over birds and dinosaurs". Science News 157 (3): 38. doi:. http://www.sciencenews.org/view/generic/id/94/title/All_mixed_up_over_birds_and_dinosaurs.
- ^ a b c d e Xu Xing; Tang Zhi-lu; & Wang Xiaolin. (1999). "A therizinosaurid dinosaur with integumentary structures from China". Nature 399 (6734): 350–354. doi:.
- ^ a b c d e f g Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q., Jia, C. (2004). "Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids". Nature 431: 680–684. doi:.
- ^ Zhou Zhonghe; & Zhang Fucheng (2002). "A long-tailed, seed-eating bird from the Early Cretaceous of China". Nature 418 (6896): 405–409. doi:.
- ^ Wellnhofer, P. (1988). Ein neuer Exemplar von Archaeopteryx. Archaeopteryx 6:1–30.
- ^ a b c Zhou Zhonghe; Barrett, Paul M.; & Hilton, Jason. (2003). "An exceptionally preserved Lower Cretaceous ecosystem". Nature 421 (6925): 807–814. doi:.
- ^ a b c d Feduccia, A., Lingham-Soliar, T. & Hinchliffe, J. R. (2005). Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence. Journal of Morphology 266, 125-166. doi:10.1002/jmor.10382
- ^ a b c Czerkas, S.A., Zhang, D., Li, J., and Li, Y. (2002). "Flying Dromaeosaurs". in Czerkas, S.J.. Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1. Blanding: The Dinosaur Museum. pp. 16–26.
- ^ a b Norell, Mark, Ji, Qiang, Gao, Keqin, Yuan, Chongxi, Zhao, Yibin, Wang, Lixia. (2002). "'Modern' feathers on a non-avian dinosaur". Nature, 416: pp. 36. 7 March 2002.
- ^ a b c d e f g h Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. ISBN 978-0801867637.
- ^ Heilmann, G. (1926): The Origin of Birds. Witherby, London. ISBN 0-486-22784-7 (1972 Dover reprint)
- ^ John Ostrom (1975). The origin of birds. Annual Review of Earth and Planetary Sciences 3, pp. 55.
- ^ Bryant, H.N. & Russell, A.P. (1993) The occurrence of clavicles within Dinosauria: implications for the homology of the avian furcula and the utility of negative evidence. Journal of Vertebrate Paleontology, 13(2):171-184.
- ^ Chure, Daniel J.; & Madsen, James H. (1996). "On the presence of furculae in some non-maniraptoran theropods". Journal of Vertebrate Paleontology 16 (3): 573–577.
- ^ Norell, Mark A.; & Makovicky, Peter J. (1999). "Important features of the dromaeosaurid skeleton II: Information from newly collected specimens of Velociraptor mongoliensis". American Museum Novitates 3282: 1–44. http://hdl.handle.net/2246/3025.
- ^ Colbert, E. H. & Morales, M. (1991) Evolution of the vertebrates: a history of the backboned animals through time. 4th ed. Wiley-Liss, New York. 470 p.
- ^ Barsbold, R. et al. (1990) Oviraptorosauria. In The Dinosauria, Weishampel, Dodson &p; Osmolska (eds) pp 249-258.
- ^ Included as a cladistic definer, e.g. (Columbia University) Master Cladograms or mentioned even in the broadest context, such as Paul C. Sereno, "The origin and evolution of dinosaurs" Annual Review of Earth and Planetary Sciences 25 pp 435-489.
- ^ Lipkin, C., Sereno, P.C., and Horner, J.R. (November 2007). "THE FURCULA IN SUCHOMIMUS TENERENSIS AND TYRANNOSAURUS REX (DINOSAURIA: THEROPODA: TETANURAE)". Journal of Paleontology 81 (6): 1523–1527. doi:. http://jpaleontol.geoscienceworld.org/cgi/content/extract/81/6/1523. - full text currently online at "The Furcula in Suchomimus Tenerensis and Tyrannosaurus rex". http://www.redorbit.com/news/health/1139122/the_furcula_in_suchomimus_tenerensis_and_tyrannosaurus_rex_dinosauria_theropoda/index.html. This lists a large number of theropods in which furculae have been found, as well as describing those of Suchomimus Tenerensis and Tyrannosaurus rex.
- ^ Carrano, M,R., Hutchinson, J.R., and Sampson, S.D. (December 2005). "New information on Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona". Journal of Vertebrate Paleontology 25 (4): 835–849. doi:. Archived from the original on 2006-05-12. http://web.archive.org/web/20060512053601/http://www.rvc.ac.uk/AboutUs/Staff/jhutchinson/documents/JH18.pdf.
- ^ Yates, Adam M.; and Vasconcelos, Cecilio C. (2005). "Furcula-like clavicles in the prosauropod dinosaur Massospondylus". Journal of Vertebrate Paleontology 25 (2): 466–468. doi:.
- ^ Downs, A. (2000). Coelophysis bauri and Syntarsus rhodesiensis compared, with comments on the preparation and preservation of fossils from the Ghost Ranch Coelophysis Quarry. New Mexico Museum of Natural History and Science Bulletin, vol. 17, pp. 33–37.
- ^ The furcula of Coelophysis bauri, a Late Triassic (Apachean) dinosaur (Theropoda: Ceratosauria) from New Mexico. 2006. By Larry Rinehart, Spencer Lucas, and Adrian Hunt
- ^ a b Ronald S. Tykoski, Catherine A. Forster, Timothy Rowe, Scott D. Sampson, and Darlington Munyikwad. (2002). A furcula in the coelophysid theropod Syntarsus. Journal of Vertebrate Paleontology 22(3):728-733.
- ^ Larry F. Rinehart, Spencer G. Lucas, Adrian P. Hunt. (2007). Furculae in the Late Triassic theropod dinosaur Coelophysis bauri. Paläontologische Zeitschrift 81: 2
- ^ a b Sereno, P.C.; Martinez, R.N.; Wilson, J.A.; Varricchio, D.J.; Alcober, O.A.; and Larsson, H.C.E. (September 2008). "Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina". PLoS ONE 3 (9): e3303. doi:. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0003303. Retrieved on 2008-10-27.
- ^ O'Connor, P.M. & Claessens, L.P.A.M. (2005). "Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs". Nature 436: 253–256. doi:.
- ^ Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System Newswise, Retrieved on September 29, 2008.
- ^ Fisher, P. E., Russell, D. A., Stoskopf, M. K., Barrick, R. E., Hammer, M. & Kuzmitz, A. A. (2000). Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur. Science 288, 503–505.
- ^ Hillenius, W. J. & Ruben, J. A. (2004). The evolution of endothermy in terrestrial vertebrates: Who? when? why? Physiological and Biochemical Zoology 77, 1019–1042.
- ^ Dinosaur with a Heart of Stone. T. Rowe, E. F. McBride, P. C. Sereno, D. A. Russell, P. E. Fisher, R. E. Barrick, and M. K. Stoskopf (2001) Science 291, 783
- ^ a b Xu, X. and Norell, M.A. (2004). A new troodontid dinosaur from China with avian-like sleeping posture. Nature 431:838-841.See commentary on the article.
- ^ Schweitzer, M.H.; Wittmeyer, J.L.; and Horner, J.R. (2005). "Gender-specific reproductive tissue in ratites and Tyrannosaurus rex". Science 308: 1456–1460. doi:. PMID 15933198. http://www.sciencemag.org/cgi/content/abstract/308/5727/1456.
- ^ Lee, Andrew H.; and Werning, Sarah (2008). "Sexual maturity in growing dinosaurs does not fit reptilian growth models". Proceedings of the National Academy of Sciences 105 (2): 582–587. doi:. PMID 18195356. http://www.pnas.org/cgi/content/abstract/105/2/582.
- ^ Chinsamy, A., Hillenius, W.J. 2004). Physiology of nonavian dinosaurs. In:Weishampel, D.B., Dodson, P., Osmolska, H., eds. The Dinosauria. University of California Press, Berkely. pp. 643-65.
- ^ Norell, M.A., Clark, J.M., Chiappe, L.M., and Dashzeveg, D. (1995). "A nesting dinosaur." Nature 378:774-776.
- ^ a b Clark, J.M., Norell, M.A., & Chiappe, L.M. (1999). "An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest." American Museum Novitates, 3265: 36 pp., 15 figs.; (American Museum of Natural History) New York. (5.4.1999).
- ^ Norell, M. A., Clark, J. M., Dashzeveg, D., Barsbold, T., Chiappe, L. M., Davidson, A. R., McKenna, M. C. and Novacek, M. J. (November 1994). "A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs" (abstract page). Science 266 (5186): 779–782. doi:. PMID 17730398. http://www.sciencemag.org/cgi/content/abstract/266/5186/779.
- ^ Oviraptor nesting Oviraptor nests or Protoceratops?
- ^ Gregory Paul (1994). Thermal environments of dinosaur nestlings: Implications for endothermy and insulation. In: Dinosaur Eggs and Babies.
- ^ Hombergerm D.G. (2002). The aerodynamically streamlined body shape of birds: Implications for the evolution of birds, feathers, and avian flight. In: Zhou, Z., Zhang, F., eds. Proceedings of the 5th symposium of the Society of Avian Paleontology and Evolution, Beijing, 1-4 June 2000. Beijing, China: Science Press. p. 227-252.
- ^ a b c Ji, Q., and Ji, S. (1997). "A Chinese archaeopterygian, Protarchaeopteryx gen. nov." Geological Science and Technology (Di Zhi Ke Ji), 238: 38-41. Translated By Will Downs Bilby Research Center Northern Arizona University January, 2001
- ^ a b c d e Xu, X., Zhou, Z., and Wang, X. (2000). "The smallest known non-avian theropod dinosaur." Nature, 408 (December): 705-708.[1]
- ^ Dal Sasso, C. and Signore, M. (1998). Exceptional soft-tissue preservation in a theropod dinosaur from Italy. Nature 292:383–387. See commentary on the article
- ^ Mary H. Schweitzer, Jennifer L. Wittmeyer, John R. Horner, and Jan K. Toporski (2005). Science 307 (5717) pp. 1952-1955. doi:10.1126/science.1108397
- ^ Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. (2005). Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307:1952–1955. See commentary on the article
- ^ Wang, H., Yan, Z. and Jin, D. (1997). Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil. Molecular Biology and Evolution. 14:589–591. See commentary on the article.
- ^ Chang, B.S.W., Jönsson, K., Kazmi, M.A., Donoghue, M.J. and Sakmar, T.P. (2002). Recreating a Functional Ancestral Archosaur Visual Pigment. Molecular Biology and Evolution 19:1483–1489. See commentary on the article.
- ^ Embery, et al. "Identification of proteinaceous material in the bone of the dinosaur Iguanodon." Connect Tissue Res. 2003; 44 Suppl 1:41-6. PMID: 12952172
- ^ Schweitzer, et al. (1997 Jun 10) "Heme compounds in dinosaur trabecular bone." Proc Natl Acad Sci U S A.. 94(12):6291–6. PMID: 9177210
- ^ Fucheng, Z., Zhonghe, Z., and Dyke, G. (2006). Feathers and 'feather-like' integumentary structures in Liaoning birds and dinosaurs. Geol . J. 41:395-404.
- ^ a b Cheng-Ming Choung, Ping Wu, Fu-Cheng Zhang, Xing Xu, Minke Yu, Randall B. Widelitz, Ting-Xin Jiang, and Lianhai Hou (2003). Adaptation to the sky: defining the feather with integument fossils from the Mesozoic China and exprimental evidence from molecular laboratories. Journal of Experimental Zoology (MOL DEV EVOL) 298b:42-56.
- ^ Bakker, R.T., Galton, P.M. (1974). Dinosaur monophyly and a new class of vertebrates. Nature 248:168-172.
- ^ Sumida, SS & CA Brochu (2000). "Phylogenetic context for the origin of feathers". American Zoologist 40 (4): 486–503. doi:. http://icb.oxfordjournals.org/cgi/content/abstract/40/4/486.
- ^ a b c d Chiappe, Luis M., (2009). Downsized Dinosaurs:The Evolutionary Transition to Modern Birds. Evo Edu Outreach 2: 248-256. doi:10.1007/s12052-009-0133-4
- ^ Burgers, P., Chiappe, L.M. (1999). The wing of Archaeopteryx as a primary thrust generator. Nature 399: 60-2. doi:10.1038/19967
- ^ a b c d e f g h Prum, R. & Brush A.H. (2002). "The evolutionary origin and diversification of feathers". The Quarterly Review of Biology 77: 261–295. doi:.
- ^ a b c d Prum, R. H. (1999). Development and evolutionary origin of feathers. Journal of Experimental Zoology 285, 291-306.
- ^ Griffiths, P. J. (2000). The evolution of feathers from dinosaur hair. Gaia 15, 399-403.
- ^ a b c d e f g Mayr, G. Peters, S.D. Plodowski, G. Vogel, O. (2002). "Bristle-like integumentary structures at the tail of the horned dinosaur Psittacosaurus". Naturwissenschaften 89: 361–365. doi:.
- ^ a b c Schweitzer, Mary Higby, Watt, J.A., Avci, R., Knapp, L., Chiappe, L, Norell, Mark A., Marshall, M. (1999). "Beta-Keratin Specific Immunological reactivity in Feather-Like Structures of the Cretaceous Alvarezsaurid, Shuvuuia deserti Journal of Experimental Zoology Part B (Mol Dev Evol) 285:146-157
- ^ Schweitzer, M. H. (2001). Evolutionary implications of possible protofeather structures associated with a specimen of Shuvuuia deserti. In Gauthier, J. & Gall, L. F. (eds) New prespectives on the origin and early evolution of birds: proceedings of the international symposium in honor of John H. Ostrom. Peabody Museum of Natural History, Yale University (New Haven), pp. 181-192.
- ^ Christiansen, P. & Bonde, N. (2004). Body plumage in Archaeopteryx: a review, and new evidence from the Berlin specimen. C. R. Palevol 3, 99-118.
- ^ M.J. Benton, M.A. Wills, R. Hitchin. (2000). Quality of the fossil record through time. Nature 403, 534-537. doi:10.1038/35000558
- ^ Morgan, James (2008-10-22). "New feathered dinosaur discovered". BBC. http://news.bbc.co.uk/2/hi/science/nature/7684796.stm. Retrieved on 2009-07-02.
- ^ a b c d e f Zhang, F., Zhou, Z., Xu, X., Wang, X., & Sullivan, C. (2008). "A bizarre Jurassic maniraptoran from China with elongate ribbon-like feathers." Available from Nature Precedings, doi:10.1038/npre.2008.2326.1 [2].
- ^ Prum, R,. O. & Brush, A. H. (2003). Which came first, the feather or the bird? Scientific American 286 (3), 84-93.
- ^ Epidexipteryx: bizarre little strap-feathered maniraptoran ScienceBlogs Tetrapod Zoology article by Darren Naish. October 23, 2008
- ^ Gishlick, A. D. (2001). The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight. In Gauthier, J. & Gall, L. F. (eds) New Perspectives on the Origin and Early Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom. Peabody Museum of Natural History, Yale University (New Haven), pp. 301-318.
- ^ Senter, P. (2006). Comparison of forelimb function between Deinonychus and Bambiraptor (Theropoda: Dromaeosauridae). Journal of Vertebrate Paleontology 26, 897-906.
- ^ JA Long, P Schouten. (2008). Feathered Dinosaurs: The Origin of Birds
- ^ a b Yalden, D. W. (1985). Forelimb function in Archaeopteryx. In Hecht, M. K., Ostrom, J. H., Viohl, G. & Wellnhofer, P. (eds) The Beginnings of Birds - Proceedings of the International Archaeopteryx Conference, Eichstatt 1984, pp. 91-97.
- ^ Chen, P.-J., Dong, Z.-M. & Zhen, S.-N. (1998). An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China. Nature 391, 147-152.
- ^ a b c Currie, Philip J.; Pei-ji Chen. (2001). Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Sciences 38, 1705-1727. doi:10.1139/cjes-38-12-1705
- ^ Bohlin, B. 1947. The wing of Archaeornithes. Zoologiska Bidrag 25, 328-334.
- ^ Rietschel, S. (1985). Feathers and wings of Archaeopteryx, and the question of her flight ability. In Hecht, M. K., Ostrom, J. H., Viohl, G. & Wellnhofer, P. (eds) The Beginnings of Birds - Proceedings of the International Archaeopteryx Conference, Eichstatt 1984, pp. 251-265.
- ^ a b Griffiths, P. J. 1993. The claws and digits of Archaeopteryx lithographica. Geobios 16, 101-106.
- ^ Stephan, B. 1994. The orientation of digital claws in birds. Journal fur Ornithologie 135, 1-16.
- ^ a b c Chiappe, L.M. and Witmer, L.M. (2002). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press, ISBN 0520200942
- ^ Martin, L. D. & Lim, J.-D. (2002). Soft body impression of the hand in Archaeopteryx. Current Science 89, 1089-1090.
- ^ a b c d Feduccia, A. (1999). The Origin and Evolution of Birds. 420 pp. Yale University Press, New Haven. ISBN 0300078617.
- ^ a b Dyke, G.J., and Norell, M.A. (2005). "Caudipteryx as a non-avialan theropod rather than a flightless bird." Acta Palaeontologica Polonica, 50(1): 101–116. PDF fulltext
- ^ a b c Witmer, L.M. (2002). “The Debate on Avian Ancestry; Phylogeny, Function and Fossils”, Mesozoic Birds: Above the Heads of Dinosaurs : 3–30. ISBN 0-520-20094-2
- ^ Jones T.D., Ruben J.A., Martin L.D., Kurochkin E.N., Feduccia A., Maderson P.F.A., Hillenius W.J., Geist N.R., Alifanov V. (2000). Nonavian feathers in a Late Triassic archosaur. Science 288: 2202-2205.
- ^ Martin, Larry D. (2006). "A basal archosaurian origin for birds". Acta Zoologica Sinica 50 (6): 977–990.
- ^ Burke, Ann C.; & Feduccia, Alan. (1997). "Developmental patterns and the identification of homologies in the avian hand". Science 278 (5338): 666–668. doi:.
- ^ a b Kevin Padian (2000). Dinosaurs and Birds — an Update. Reports of the National Center for Science Education. 20 (5):28–31.
- ^ Ostrom J.H. (1973). The ancestry of birds. Nature 242: 136.
- ^ a b Padian, Kevin. (2004). "Basal Avialae". in Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 210–231. ISBN 0-520-24209-2.
- ^ Olshevsky, G. (1991). A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia. Publications Requiring Research, San Diego.
- ^ Olshevsky, G. (1994). The birds first? A theory to fit the facts. Omni 16 (9), 34-86.
- ^ a b Chatterjee, S. (1999): Protoavis and the early evolution of birds. Palaeontographica A 254: 1-100.
- ^ Chatterjee, S. (1995): The Triassic bird Protoavis. Archaeopteryx 13: 15-31.
- ^ Chatterjee, S. (1998): The avian status of Protoavis. Archaeopteryx 16: 99-122.
- ^ Chatterjee, S. (1991). "Cranial anatomy and relationships of a new Triassic bird from Texas." Philosophical Transactions of the Royal Society B: Biological Sciences, 332: 277-342. HTML abstract
- ^ Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore. ISBN 0-8018-6763-0
- ^ Witmer, L. (2002). "The debate on avian ancestry: phylogeny, function, and fossils." Pp. 3-30 in: Chiappe, L.M. and Witmer, L.M. (eds), Mesozoic birds: Above the heads of dinosaurs. University of California Press, Berkeley, Calif., USA. ISBN 0-520-20094-2
- ^ Nesbitt, Sterling J.; Irmis, Randall B. & Parker, William G. (2007): A critical re-evaluation of the Late Triassic dinosaur taxa of North America. Journal of Systematic Palaeontology 5(2): 209-243.
- ^ Ostrom, J. (1987): Protoavis, a Triassic bird? Archaeopteryx 5: 113-114.
- ^ Ostrom, J.H. (1991): The bird in the bush. Nature 353(6341): 212.
- ^ Ostrom, J.H. (1996): The questionable validity of Protoavis. Archaeopteryx 14: 39-42.
- ^ Chatterjee, S. (1987). "Skull of Protoavis and Early Evolution of Birds." Journal of Vertebrate Paleontology, 7(3)(Suppl.): 14A.
- ^ a b EvoWiki (2004). Chatterjee's Chimera: A Cold Look at the Protoavis Controversy. Version of 2007-JAN-22. Retrieved 2009-FEB-04.
- ^ Chatterjee, S. (1997). The Rise of Birds: 225 Million Years of Evolution. Johns Hopkins University Press, Baltimore. ISBN 0-8018-5615-9
- ^ Feduccia, Alan (1994) "The Great Dinosaur Debate" Living Bird. 13:29-33.
- ^ Why Birds Aren't Dinosaurs. Explore:Thought and Discovery at the University of Kansas. Accessed 8/05/09.
- ^ Jensen, James A. & Padian, Kevin. (1989) "Small pterosaurs and dinosaurs from the Uncompahgre fauna (Brushy Basin member, Morrison Formation: ?Tithonian), Late Jurassic, western Colorado" Journal of Paleontology Vol. 63 no. 3 pg. 364-373.
- ^ Lubbe, T. van der, Richter, U., and Knötschke, N. 2009. Velociraptorine dromaeosaurid teeth from the Kimmeridgian (Late Jurassic) of Germany. Acta Palaeontologica Polonica 54 (3): 401–408. DOI: 10.4202/app.2008.0007.
- ^ a b c d Hartman, S., Lovelace, D., and Wahl, W., (2005). "Phylogenetic assessment of a maniraptoran from the Morrison Formation." Journal of Vertebrate Paleontology, 25, Supplement to No. 3, pp 67A-68A http://www.bhbfonline.org/AboutUs/Lori.pdf
- ^ Brochu, Christopher A. Norell, Mark A. (2001) "Time and trees: A quantitative assessment of temporal congruence in the bird origins debate" pp.511-535 in "New Perspectives on the Origin and Early Evolution of Birds" Gauthier&Gall, ed. Yale Peabody Museum. New Haven, Conn. USA.
- ^ a b Ruben, J., Jones, T. D., Geist, N. R. & Hillenius, W. J. (1997). Lung structure and ventilation in theropod dinosaurs and early birds. Science 278, 1267-1270.
- ^ a b Ruben, J., Dal Sasso, C., Geist, N. R., Hillenius, W. J., Jones, T. D. & Signore, M. (1999). Pulmonary function and metabolic physiology of theropod dinosaurs. Science 283, 514-516.
- ^ Quick, D. E. & Ruben, J. A. (2009). Cardio-pulmonary anatomy in theropod dinosaurs: implications from extant archosaurs. Journal of Morphology doi: 10.1002/jmor.10752
- ^ gazettetimes.com article
- ^ Discovery Raises New Doubts About Dinosaur-bird Links ScienceDaily article
- ^ Ruben, J., Hillenius, W., Geist, N. R., Leitch, A., Jones, T. D., Currie, P. J., Horner, J. R. & Espe, G. (1996). The metabolic status of some Late Cretaceous dinosaurs. Science 273, 1204-1207.
- ^ Theagarten Lingham-Soliar (2003). The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers. Naturwissenschaften 90 (12): 563-567.
- ^ Peter Wellnhofer (2004) "Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Chapter 13. The Plumage of Archaeopteryx:Feathers of a Dinosaur?" Currie, Koppelhaus, Shugar, Wright. Indiana University Press. Bloomington, IN. USA. pp. 282-300.
- ^ Lingham-Soliar, T et al. (2007) Proc. R. Soc. Lond. B doi:10.1098/rspb.2007.0352.
- ^ Access : Bald dino casts doubt on feather theory : Nature News
- ^ "Transcript: The Dinosaur that Fooled the World". BBC. http://www.bbc.co.uk/science/horizon/2001/dinofooltrans.shtml. Retrieved on 2006-12-22.
- ^ Mayell, Hillary (2002-11-20). "Dino Hoax Was Mainly Made of Ancient Bird, Study Says". National Geographic. http://news.nationalgeographic.com/news/2002/11/1120_021120_raptor.html. Retrieved on 2008-06-13.
- ^ Zhou, Zhonghe, Clarke, Julia A., Zhang, Fucheng. "Archaeoraptor's better half." Nature Vol. 420. 21 November 2002. pp. 285.
- ^ a b Makovicky, Peter J.; Apesteguía, Sebastián; & Agnolín, Federico L. (2005). "The earliest dromaeosaurid theropod from South America". Nature 437 (7061): 1007–1011. doi:.
- ^ Norell, M.A., Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. (2006). "A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia)." American Museum Novitates, 3545: 1-51.
- ^ a b Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M. & Krause, David W. (1998). "The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar". Science (5358): pp. 1915–1919. doi:10.1126/science.279.5358.1915. (HTML abstract).
- ^ a b Chiappe, L.M.. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Sydney: UNSW Press.
- ^ a b Kurochkin, E., N. (2006). Parallel Evolution of Theropod Dinoaurs and Birds. Entomological Review 86 (1), pp. S45-S58. doi:10.1134/S0013873806100046
- ^ Kurochkin, E., N. (2004). A Four-Winged Dinosaur and the Origin of Birds. Priroda 5, 3-12.
- ^ a b c S. Chatterjee. (2005). The Feathered Dinosaur Microraptor:Its Biplane Wing Platform and Flight Performance. 2005 Salt Lake City Annual Meeting.
- ^ a b c d Chatterjee, S., and Templin, R.J. (2007). "Biplane wing platform and flight performance of the feathered dinosaur Microraptor gui." Proceedings of the National Academy of Sciences, 104(5): 1576-1580. [3]
- ^ a b c Holtz, Thomas R.; & Osmólska, Halszka. (2004). "Saurischia". in Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 21–24. ISBN 0-520-24209-2.
- ^ a b c d e Xu, Xing; Zheng Xiao-ting; You, Hai-lu (20 January 2009). "A new feather type in a nonavian theropod and the early evolution of feathers". Proceedings of the National Academy of Sciences (Philadelphia). doi:10.1073/pnas.0810055106. PMID 19139401.
- ^ a b c Turner, Alan H.; Hwang, Sunny; & Norell, Mark A. (2007). "A small derived theropod from Öösh, Early Cretaceous, Baykhangor, Mongolia". American Museum Novitates 3557 (3557): 1–27. doi:. http://hdl.handle.net/2246/5845.
- ^ a b Bryner, Jeanna (2009). "Ancient Dinosaur Wore Primitive Down Coat." http://www.foxnews.com/story/0,2933,479875,00.html
- ^ a b Xu, X., Cheng, C., Wang, X. & Chang, C. (2003). Pygostyle-like structure from Beipiaosaurus (Theropoda, Therizinosauroidea) from the Lower Cretaceous Yixian Formation of Liaoning, China. Acta Geologica Sinica 77, 294-298.
- ^ a b c Xu Xing; Zhou Zhonghe & Prum, Richard A. (2003). "Branched integumental structures in Sinornithosaurus and the origin of feathers". Nature 410 (6825): 200–204. doi:.
- ^ Paul, Gregory S. (2008). "The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia". in Carpenter, Kenneth; and Larson, Peter E. (editors). Tyrannosaurus rex, the Tyrant King (Life of the Past). Bloomington: Indiana University Press. p. 316. ISBN 0-253-35087-5.
- ^ Martin, Larry D.; & Czerkas, Stephan A. (2000). "The fossil record of feather evolution in the Mesozoic". American Zoologist 40 (4): 687–694. doi:. http://www.bioone.org/perlserv/?request=get-abstract&doi=10.1668%2F0003-1569%282000%29040%5B0687%3ATFROFE%5D2.0.CO%3B2.
- ^ a b T. rex was fierce, yes, but feathered, too.
- ^ Nicholas M. Gardner, David B. Baum, Susan Offner. (2008). [392b%3ANDEFFI2.0.CO%3B2 No Direct Evidence for Feathers in Tyrannosaurus rex]. The American Biology Teacher 70(7):392-392
- ^ a b Xu, X., Wang, X.-L., and Wu, X.-C. (1999). "A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China". Nature 401: 262–266. doi:.
- ^ a b c d e f g h i j k Turner, A.H.; Makovicky, P.J.; and Norell, M.A. (2007). "Feather quill knobs in the dinosaur Velociraptor" (pdf). Science 317 (5845): 1721. doi:. PMID 17885130. http://www.sciencemag.org/cgi/reprint/317/5845/1721.pdf.
- ^ a b Ji, Q., Norell, M. A., Gao, K.-Q., Ji, S.-A. & Ren, D. (2001). The distribution of integumentary structures in a feathered dinosaur. Nature 410, 1084-1088.
- ^ a b American Museum of Natural History. "Velociraptor Had Feathers." ScienceDaily 20 September 2007. 23 January 2008 http://www.sciencedaily.com/releases/2007/09/070920145402.htm
- ^ a b c d e f g h Xu, X., and Zhang, F. (2005). "A new maniraptoran dinosaur from China with long feathers on the metatarsus." Naturwissenschaften, 92(4): 173 - 177.
- ^ a b c d Xu, X., Zhao, Q., Norell, M., Sullivan, C., Hone, D., Erickson, G., Wang, X., Han, F. and Guo, Y. (2009). "A new feathered maniraptoran dinosaur fossil that fills a morphological gap in avian origin." Chinese Science Bulletin, 6 pages, accepted November 15, 2008.
- ^ [4] Currie, PJ & Chen, PJ (2001) Anatomy of Sinosauropteryx prima from Liaoning, northeastern China, Canadian Journal of Earth Sciences, 38: 1,705-1,727.
- ^ Buffetaut, E., Grellet-Tinner, G., Suteethorn, V., Cuny, G., Tong, H., Košir, A., Cavin, L., Chitsing, S., Griffiths, P.J., Tabouelle, J. and Le Loeuff, J. (2005). "Minute theropod eggs and embryo from the Lower Cretaceous of Thailand and the dinosaur-bird transition." Naturwissenschaften, 92(10): 477-482.
- ^ a b c Czerkas, S.A., and Yuan, C. (2002). "An arboreal maniraptoran from northeast China." Pp. 63-95 in Czerkas, S.J. (Ed.), Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal 1. The Dinosaur Museum, Blanding, U.S.A. PDF abridged version
- ^ Maryanska, T., Osmolska, H., & Wolsam, M. (2002). "Avialian status for Oviraptorosauria". Acta Palaeontologica Polonica 47 (1): 97–116.
- ^ Benton, M. J. (2004). Vertebrate Palaeontology, 3rd ed. Blackwell Science Ltd.
- ^ a b Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; and Norell, Mark (2007). "A basal dromaeosaurid and size evolution preceding avian flight" (pdf). Science 317: 1378–1381. doi:. PMID 17823350. http://www.sciencemag.org/cgi/reprint/317/5843/1378.pdf.
- ^ a b c d Barsbold, R., Osmólska, H., Watabe, M., Currie, P.J., and Tsogtbaatar, K. (2000). "New Oviraptorosaur (Dinosauria, Theropoda) From Mongolia: The First Dinosaur With A Pygostyle". Acta Palaeontologica Polonica, 45(2): 97-106.
- ^ C.M. Chuong, R. Chodankar, R.B. Widelitz (2000). Evo-Devo of feathers and scales: building complex epithelial appendages. Commentary, Current Opinion in Genetics & Development 10 (4), pp. 449-456.
- ^ a b Kurzanov, S.M. (1987). "Avimimidae and the problem of the origin of birds." Transactions of the Joint Soviet-Mongolian Paleontological Expedition, 31: 5-92. [in Russian]
- ^ a b Hopp, Thomas J., Orsen, Mark J. (2004) "Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Chapter 11. Dinosaur Brooding Behavior and the Origin of Flight Feathers" Currie, Koppelhaus, Shugar, Wright. Indiana University Press. Bloomington, IN. USA.
- ^ Maryańska, T. & Osmólska, H. (1997). The Quadrate of Oviraptorid Dinosaurs. Acta Palaeontologica Polonica 42 (3): 361-371.
- ^ Jones, T.D., Farlow, J.O., Ruben, J.A., Henderson, D.M., and Hillenius, W.J. (2000). "Cursoriality in bipedal archosaurs." Nature, 406(6797): 716–718. doi:10.1038/35021041 PDF fulltext Supplementary information
- ^ Zhou, Z., Wang, X., Zhang, F., and Xu, X. (2000). "Important features of Caudipteryx - Evidence from two nearly complete new specimens." Vertebrata Palasiatica, 38(4): 241–254. PDF fulltext
- ^ Buchholz, P. (1997). Pelecanimimus polyodon. Dinosaur Discoveries 3, 3-4.
- ^ Briggs, D. E., Wilby, P. R., Perez-Moreno, B., Sanz, J. L. & Fregenal-Martinez, M. (1997). The mineralization of dinosaur soft tissue in the Lower Cretaceous of Las Hoyas, Spain. Journal of the Geological Society, London 154, 587-588.
- ^ a b Theagarten Lingham-Soliar. (2008). A unique cross section through the skin of the dinosaur Psittacosaurus from China showing a complex fibre architecture. Proc R Soc B 275: 775-780.
- ^ a b c d e f g h i j k Zheng, X.-T., You, H.-L., Xu, X. and Dong, Z.-M. (2009). "An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures." Nature, 458(19): 333-336. doi:10.1038/nature07856
- ^ Witmer, L.M. (2009), "Dinosaurs: Fuzzy origins for feathers", Nature 458 (7236): 293–295, http://www.nature.com/nature/journal/v458/n7236/full/458293a.html, retrieved on 2009-09-02
- ^ "Tianyulong". Pharyngula. PZ Myers. March 20, 2009. http://scienceblogs.com/pharyngula/2009/03/tianyulong.php. Retrieved on 2009-04-30.
- ^ a b "Tianyulong - a fuzzy dinosaur that makes the origin of feathers fuzzier". Not Exactly Rocket Science:Science for Everyone. Ed Yong. March 18, 2009. http://scienceblogs.com/notrocketscience/2009/03/tianyulong_-_a_fuzzy_dinosaur_that_makes_the_origin_of_feath.php. Retrieved on 2009-07-22.
- ^ Xu, X., Wang, X., Wu, X., (1999). A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China. Nature 401:6750 262-266 doi 10.1038/45769
- ^ Xu. X., Zhao, X., Clark, J.M., (1999). A new therizinosaur from the Lower Jurassic lower Lufeng Formation of Yunnan, China. Journal of Vertebrate Paleontology 21:3 477–483 doi 10.1671/0272-4634
- ^ Xu, X. and Wang, X.-L. (2003). "A new maniraptoran from the Early Cretaceous Yixian Formation of western Liaoning." Vertebrata PalAsiatica, 41(3): 195–202.
- ^ Ji, Q., Ji, S., Lu, J., You, H., Chen, W., Liu, Y., and Liu, Y. (2005). "First avialan bird from China (Jinfengopteryx elegans gen. et sp. nov.)." Geological Bulletin of China, 24(3): 197-205.
- ^ Ji, S., Ji, Q., Lu J., and Yuan, C. (2007). "A new giant compsognathid dinosaur with long filamentous integuments from Lower Cretaceous of Northeastern China." Acta Geologica Sinica, 81(1): 8-15.
- ^ Czerkas, S.A., and Ji, Q. (2002). "A new rhamphorhynchoid with a headcrest and complex integumentary structures." Pp. 15-41 in: Czerkas, S.J. (Ed.). Feathered Dinosaurs and the Origin of Flight. Blanding, Utah: The Dinosaur Museum. ISBN 1-93207-501-1.
- ^ a b c Senter, Phil (2007). "A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)". Journal of Systematic Palaeontology 5 (4): 429–463. doi:.
- ^ Osmólska, Halszka; Maryańska, Teresa; & Wolsan, Mieczysław. (2002). "Avialan status for Oviraptorosauria". Acta Palaeontologica Polonica 47 (1): 97–116. http://app.pan.pl/article/item/app47-097.html.
- ^ Martinelli, Agustín G.; & Vera, Ezequiel I. (2007). "Achillesaurus manazzonei, a new alvarezsaurid theropod (Dinosauria) from the Late Cretaceous Bajo de la Carpa Formation, Río Negro Province, Argentina". Zootaxa 1582: 1–17. http://www.mapress.com/zootaxa/2007f/z01582p017f.pdf.
- ^ Novas, Fernando E.; & Pol, Diego. (2002). "Alvarezsaurid relationships reconsidered". in Chiappe, Luis M.; & Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. pp. 121–125. ISBN 0-520-20094-2.
- ^ Sereno, Paul C. (1999). "The evolution of dinosaurs". Science 284 (5423): 2137–2147. doi:. PMID 10381873.
- ^ Perle, Altangerel; Norell, Mark A.; Chiappe, Luis M.; & Clark, James M. (1993). "Flightless bird from the Cretaceous of Mongolia". Science 362 (6421): 623–626. doi:.
- ^ Chiappe, Luis M.; Norell, Mark A.; & Clark, James M. (2002). "The Cretaceous, short-armed Alvarezsauridae: Mononykus and its kin". in Chiappe, Luis M.; & Witmer, Lawrence M. (eds.). Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. pp. 87–119. ISBN 0-520-20094-2.
- ^ Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M.; & Krause, David W. (1998). "The theropod ancestry of birds: new evidence from the Late Cretaceous of Madagascar". Science 279 (5358): 1915–1919. doi:. PMID 9506938.
- ^ Mayr, Gerald; Pohl, Burkhard; & Peters, D. Stefan (2005). "A well-preserved Archaeopteryx specimen with theropod features.". Science 310 (5753): 1483–1486. doi:. PMID 16322455.
- ^ Göhlich, U.B., and Chiappe, L.M. (2006). "A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago." Nature, 440: 329-332.
Further reading[]
- Gauthier, J.; De Queiroz, K. (2001), "Feathered dinosaurs, flying dinosaurs, crown dinosaurs, and the name" Aves", New Perspectives on the Origin and Early Evolution of Birds: 7–41.
- Fucheng, Z.; Zhonghe, Z.; Dyke, G. (2006), "Feathers and'feather-like'integumentary structures in Liaoning birds and dinosaurs", Geological Journal 41.
- Zhou, Z. (2004), "The origin and early evolution of birds: discoveries, disputes, and perspectives from fossil evidence", Naturwissenschaften 91 (10): 455–471.
- Vargas, A.O.; Fallon, J.F. (2005), "Birds have dinosaur wings: the molecular evidence", J Exp Zool (Mol Dev Evol) 304: 86–90.
- Prum, R.O. (2002), "Why ornithologists should care about the theropod origin of birds", The Auk 119 (1): 1–17.
- Clark, J.M.; Norell, M.A.; Makovicky, P.J. (2002). "Cladistic approaches to the relationships of birds to other theropod dinosaurs". Mesozoic birds, above the heads of the dinosaurs. pp. 31–61.
- Perrichot, V.; Marion, L.; Néraudeau, D.; Vullo, R.; Tafforeau, P. (2008), "The early evolution of feathers: fossil evidence from Cretaceous amber of France", Proceedings of the Royal Society B: Biological Sciences 275 (1639): 1197.
External links[]
- DinoBuzz — dinosaur-bird controversy explained, by UC Berkeley.
- Journal of Dinosaur Paleontology, with many articles on dinosaur-bird links.
- Feathered dinosaurs at the American Museum of Natural History.
- First Dinosaur Found With its Body Covering Intact; Displays Primitive Feathers From Head to Tail — AMNH Press Release
- Notes from recent papers on theropod dinosaurs and early avians
- The evolution of feathers