| Cloudinidae Fossil range: Ediacaran | |
|---|---|
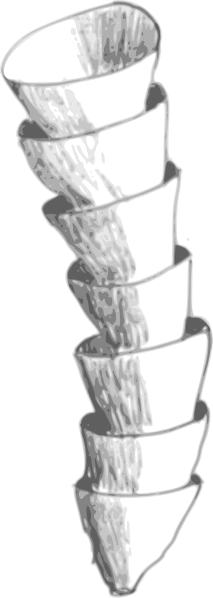 Artist's impression of a Cloudina fossil. | |
| Scientific classification
| |
|
Archaeocyatha? | |
|
Cribriocyathea? | |
|
Pterocyathida? | |
|
Cloudinidae | |
|
Cloudina | |
| |
Cloudinidae, (plural:Cloudinids) is an early metazoan family containing the genus Cloudina, which lived during the Late Ediacaran period and became extinct at the base of the Cambrian. They formed millimeter-scale conical fossils consisting of calcareous cones nested within one another; the appearance of the organism itself remains unknown. The name Cloudina honors the 20th century geologist and paleontologist Preston Cloud.[1]
Cloudinids had a wide geographic range, reflected in the present distribution of localities in which their fossils are found, and are an abundant component of some deposits. They never appear in the same layers as soft-bodied Ediacaran biota, but the fact that some sequences contain Cloudinids and Ediacaran biota in alternating layers suggests that these groups had different environmental preferences. It has been suggested that Cloudinids lived embedded in microbial mats, growing new cones to avoid being buried by silt. However no specimens have been found embedded in mats, and their mode of life is still an unresolved question.
The classification of the Cloudinids has proved difficult: they were initially regarded as polychaete worms, and then as coral-like cnidarians on the basis of what look like buds on some specimens. Current scientific opinion is divided between classifying them as polychaetes and regarding it as unsafe to classify them as members of any broader grouping.
Cloudinids are important in the history of animal evolution for two reasons. They are among the earliest and most abundant of the small shelly fossils with mineralized skeletons, and therefore feature in the debate about why such skeletons first appeared in the Late Ediacaran. The most widely-supported answer is that their shells are a defense against predators, as some specimens bear the marks of multiple attacks, which suggests they survived at least a few of them. The holes made by predators are approximately proportional to the size of the Cloudina specimens, and Sinotubulites fossils, which are often found in the same beds, have so far shown no such holes. These two points suggest that predators attacked in a selective manner, and the evolutionary arms race which this indicates is commonly cited as a cause of the Cambrian explosion of animal diversity and complexity.
Morphology[]

{kind=link}
Cutaway diagram of Cloudina showing "living space" within the shell.
Cloudina varies in size from a diameter of 0.3 to 6.5 mm, and 8 to 150 mm in length.[1] Fossils consist of a series of stacked vase-like calcite tubes, whose original mineral composition is unknown.[2] Each cone traps a significant pore space beneath it, and stacks eccentrically in the one below. This results in a ridged external appearance. The overall tube is curved or sinuous, and occasionally branches. The tube walls are 8 to 50 micrometers thick, usually lying in the range 10 to 25 μm.[3] Although it used to be thought that the tubes had test-tube like bases,[1] detailed three-dimensional reconstruction has shown that the tubes had an open base.[4] There is evidence that the tube was flexible.[5]
Classification[]
Cloudina was originally classified in 1972 as a member of the Cribricyathea, a class known from the Early Cambrian.[1] Glaessner (1976) accepted this classification and also proposed that Cloudina was similar to the annelid worms, particularly serpulid polychaetes.[6] However, Hahn & Pflug (1985) and Conway Morris et al.. (1990) doubted both Germs’ and Glaessner’s suggested relationships, and were unwilling to classify it to anything more than its own family, Cloudinidae.[7][8] Some specimens of Cloudina hartmannae display budding,[1] which implies asexual reproduction.[9] On this basis Grant (1990) classified Cloudina as a coral-like cnidarian.[3] Since the tubes had an open base, creating a single living space rather than a series of separate chambers, Cloudina is more likely to be a stem group polychaete worm,[4] in other words an evolutionary "aunt" or "cousin" of more recent polychaetes. This interpretation is reinforced by the even distribution of bore-holes made by predators.[10][11] However, as with so many Ediacaran life forms, there is great debate surrounding its position in the tree of life, and classification between the kingdom and family level may be unwise.[3][8]
Ecology[]
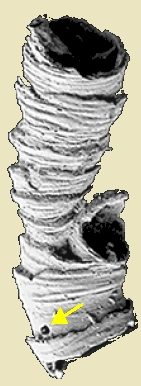
{kind=link}
Specimen showing budding structure, and predatory boring (arrow).
Cloudina is usually found in association with microbial stromatolites, which are limited to shallow water; their isotopic composition[12] suggests that water temperatures were relatively cool. They have also been found in normal sea-floor sediments, suggesting that they were not only restricted to dwelling on microbial mounds.[13] On the other hand Cloudina has never been found in the same layers as the soft-bodied Ediacara biota, but Cloudina and Ediacara biota have been found in alternating layers. This suggests that the two groups of organisms had different environmental preferences.[4]
In many Cloudina specimens the ridges formed by the cones are of varying width, which suggests the organisms grew at a variable rate. Adolf Seilacher suggests that they adhered to microbial mats, and that the growth phases represented the organism keeping pace with sedimentation - growing through new material deposited on it that would otherwise bury it. Kinks in the developing tube are easily explained by the mat falling slightly from the horizontal.[14] Because of its small size, Cloudina would be expected to be found in situ in the microbial mat, especially if, as Seilacher suggests, sedimentation built up around it during its lifetime. But all the many specimens discovered to date have only been found having been washed out of their places of growth. A further argument against Seilacher's hypothesis is that the predatory borings found in many specimens are not concentrated at what would be the top end, as one would expect if the animal was mainly buried. An alternative is that the organism dwelt on seaweeds,[4] but until a specimen unquestionably in situ is discovered, its mode of life remains open to debate.
The tubes often appear to form colonies, although they are sometimes found in more isolated situations. The frequent appearance of large and sometimes single-species colonies has be attributed to the lack of significant predation.[1] On the other hand, in some locations up to 20% of Cloudina fossils contain predatory borings ranging from 15 to 400 µm in diameter.[10][11] The boreholes are rather evenly distributed along the tube length, and some tubes had been bored multiple times — hence the organism could survive attacks, since predators do not attack empty shells. This may indicate that the animal could vary its position in the tube in response to predation, or that it occupied the full length - but not the full width - of the tube. The even distribution is perhaps difficult to reconcile with an infaunal lifestyle, mainly buried in a microbial mat, and adds weight to Miller's suggestion that the animal lived on seaweeds or in a reef environment. If modern-day molluscs are a suitable analogy, the size distribution of the borings suggests that the predator was similar in size to Cloudina.[5]
Fossil locations[]
Cloudina occurred in calcium carbonate rich areas of stromatolite reefs. It is found in association with Namacalathus, which like Cloudina was "weakly skeletal" and solitary, and Namapoikia, which was "robustly skeletal" and formed sheets on open surfaces.[15]
First found in the Nama Formation in Namibia,[1] Cloudina has also been reported in Oman,[8] China's Dengying Formation,[8][11] Canada,[16] Uruguay,[17][18] Argentina,[19] Antarctica,[20] Brazil,[21] Nevada,[22] central Spain, northwest Mexico and California.[3] It is never found in association with fossils of Cambrian age,[23] although it is never in the same bed as "Vendozoan-type" (soft-bodied) Ediacaran fossils, it is sometimes interbedded with them, suggesting that the two classes of fossil may have occupied different habitats in close proximity.
This co-existence stands against Mark McMenamin's hypothesis that the advent of skeletonisation caused the extinction of soft-bodied Ediacaran forms; however the disappearance of Cloudina at the Ediacaran - Cambrian boundary suggests that an extinction event may have occurred, which may have been related to the subsequent Cambrian explosion of life-forms. Although no Cloudinid body fossils have been found after the Ediacaran period, it has been suggested that the enigmatic trace fossils Salterella and Cornulites were formed by Cloudinids.
Paleontological importance[]
Although not the first small shelly fossil to be found, Cloudina is one of the earliest and most abundant.[24] The evolution of external shells in the Late Ediacaran is thought to be a defence against predators, marking the start of an evolutionary arms race.[24][25] While predatory borings are common in Cloudina speciments, no such borings have been found in Sinotubulites, a similar shelly fossil sometimes found in the same beds. In addition, the diameters of borings in Cloudina are proportional to the sizes of specimens, suggests that predators were selective about the size of their prey. These two indications that predators attacked selectively suggest the possibility of speciation in response to predation, which is often postulated as a potential cause of the rapid diversification of animals in the Early Cambrian.[11]
The fact that beds containing Cloudina and other shelly fossils interleave with beds containing soft-bodied Ediacara biota casts doubt on McMenamin's hypothesis that the advent of skeletonization caused the extinction of soft-bodied Ediacaran forms.[26] However there is evidence of an end-Ediacaran extinction event, which may account for the disappearance of both Cloudina and most of the Ediacara biota, and which may have been related to the subsequent Cambrian explosion of life-forms.[27]
References[]
- ^ a b c d e f g Germs, G.J.B. (October 1972). "New shelly fossils from Nama Group, South West Africa". American Journal of Science 272: 752–761.
- ^ Porter, S.M. (1 June 2007). "Seawater Chemistry and Early Carbonate Biomineralization". Science 316 (5829): 1302. doi:. PMID 17540895.
- ^ a b c d Grant, S.W. (1990). "Shell structure and distribution of Cloudina, a potential index fossil for the terminal Proterozoic". American Journal of Science (290-A): 261–294. http://lib.bioinfo.pl/pmid:11538690. Retrieved on 2008-07-19.
- ^ a b c d Miller, A.J. (2004) (PDF), A Revised Morphology of Cloudina with Ecological and Phylogenetic Implications, http://ajm.pioneeringprojects.org/files/CloudinaPaper_Final.pdf, retrieved on 2007-04-24
- ^ a b Brain, CK (2001). "Some observations on Cloudina, a terminal Proterozoic index fossil from Namibia". Journal of African Earth Sciences 33 (3): 475–480. doi:.
- ^ Glaessner, M. F (1976). "Early Phanerozoic annelid worms and their geological and biological significance". Journal of the Geological Society (London) 132: 259–275. doi:.
- ^ Hahn, G., and H. D. Pflug (1985). "Die Cloudinidae n. fam., Kalk-Röhren aus dem Vendium und Unter-Kambrium". Senckenbergiana lethaea 65: 413–431.
- ^ a b c d Conway Morris, S.; Mattes, B.W., Chen, M. (1990). "The early skeletal organism Cloudina: new occurrences from Oman and possibly China". American Journal of Science 290: 245–260.
- ^ Hua, H.; Chen, Z., Yuan, X., Zhang, L., Xiao, S. (2005). "Skeletogenesis and asexual reproduction in the earliest biomineralizing animal Cloudina". Geology 33 (4): 277–280. doi:. http://geology.geoscienceworld.org/cgi/content/abstract/33/4/277. Retrieved on 2007-04-24.
- ^ a b Hua, H.; Pratt, B.R., Zhang, L.U.Y.I. (2003). "Borings in Cloudina Shells: Complex Predator-Prey Dynamics in the Terminal Neoproterozoic". Palaios 18: 454. doi:.
- ^ a b c d Bengtson, S.; Zhao, Y. (1992-07-17). "Predatorial Borings in Late Precambrian Mineralized Exoskeletons". Science 257 (5068): 367. doi:. PMID 17832833. http://www.sciencemag.org/cgi/content/abstract/257/5068/367. Retrieved on 2007-04-24.
- ^ Ca/Mg ratios
- ^ Domke, Kirk L.; Bottjer, David J.; Loyd, Sean J.; Corsetti, Frank A.; Lyons, Timothy W. (August 2009). "Providing a Palaeoecological and Geochemical Context for Cloudina in Western North America". in Smith, Martin R.; O'Brien, Lorna J.; Caron, Jean-Bernard. International Conference on the Cambrian Explosion (Walcott 2009). Abstract Volume. Toronto, Ontario, Canada: The Burgess Shale Consortium. 31st July 2009. ISBN 978-0-9812885-1-2. http://burgess-shale.info/abstract/domke.
- ^ Seilacher, A. (1999). "Biomat-related lifestyles in the Precambrian". Palaios 14 (1): 86–93. doi:.
- ^ Neoproterozoic Microbial-Metazoan Reefs, Nama Region, Namibia - abstract retrieved January 13, 2007
- ^ Hofmann, H.J.; Mountjoy, E.W. (2001-12-01). "Namacalathus-Cloudina assemblage in Neoproterozoic Miette Group (Byng Formation), British Columbia: Canada's oldest shelly fossils". Geology 29 (12): 1091–1094. doi:. http://geology.geoscienceworld.org/cgi/content/abstract/29/12/1091. Retrieved on 2007-05-18.
- ^ Gaucher, C.; Sprechmann, P. (1998). "Grupo Arroyo del Soldado: paleontologia, edad y correlaciones (Vendiano-Cámbrico Inferior, Uruguay)" (in Spanish). Actas II Congreso Uruguaya de Geologia, Montevideo, Sociedad Uruguaya de Geologia — Facultad de Ciencias: 183–187.
- ^ Gaucher, C. (2000). Sedimentology, palaeontology, and stratigraphy of the Arroyo del Soldado Group (Vendian to Cambrian, Uruguay).
- ^ by Yochelson and Herrera, 1974; they could have mistaken them for Salterella. See Grant 1990 for reference and discussion.
- ^ Yochelson, E.L.; Stump, E. (1977). "Discovery of Early Cambrian Fossils at Taylor Nunatak, Antarctica". Journal of Paleontology 51 (4): 872–875. http://links.jstor.org/sici?sici=0022-3360(197707)51%3A4%3C872%3ADOECFA%3E2.0.CO%3B2-G. Retrieved on 2007-05-18.
- ^ Zaine, M.F.; Fairchild, T.R. (1985). "Comparison of Aulophycus lucianoi Beurlen & Sommer from Ladario (MS) and the genus Cloudina Germs, Ediacaran of Namibia". Anais Academia Brasileira de Ciencias 57: 130.
- ^ Hagadorn, J.W.; Waggoner, B. (2000). "Ediacaran fossils from the southwestern Great Basin, United States". Journal of Paleontology 74:349-359. 74: 349. doi:. http://jpaleontol.geoscienceworld.org/cgi/content/abstract/74/2/349.
- ^ Amthor, J.E.; Grotzinger, J.P., Schroder, S., Bowring, S.A., Ramezani, J., Martin, M.W., Matter, A. (2003). "Extinction of Cloudina and Namacalathus at the Precambrian-Cambrian boundary in Oman". Geology 31 (5): 431–434. doi:.
- ^ a b Bengtson, S. (2004), Early skeletal fossils, in Lipps, J.H., and Waggoner, B.M., "Neoproterozoic- Cambrian Biological Revolutions" (PDF), Palentological Society Papers 10: 67–78, http://www.cosmonova.org/download/18.4e32c81078a8d9249800021554/Bengtson2004ESF.pdf, retrieved on 2008-07-18
- ^ Dzik, J. (2007), "The Verdun Syndrome: simultaneous origin of protective armour and infaunal shelters at the Precambrian–Cambrian Transition" (PDF), Geological Society, London, Special Publications 286: 405–414;, doi:, http://www.paleo.pan.pl/people/Dzik/Publications/Verdun.pdf, retrieved on 2008-07-30
- ^ McMenamin M. (1986). The Garden of Ediacara. New York: Columbia Univ Press. ISBN 9780231105590.
- ^ Brasier, M., and Antcliffe, J. (20 August 2004). "Decoding the Ediacaran Enigma". Science 305 (5687): 1115. doi:. PMID 15326344. http://www.sciencemag.org/cgi/content/summary/305/5687/1115?ck=nck. Retrieved on 2008-07-18.